

Recruitment of endophilin to clathrin-coated pit necks is required for efficient vesicle uncoating after fission
- PMID: 22099461
- PMCID: PMC3258500
- DOI: 10.1016/j.neuron.2011.08.029
Recruitment of endophilin to clathrin-coated pit necks is required for efficient vesicle uncoating after fission
Abstract
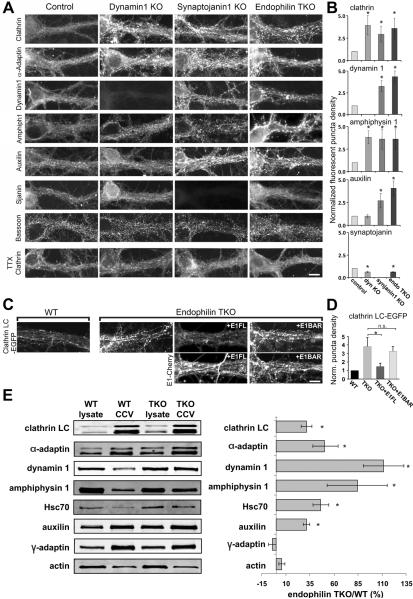
Endophilin is a membrane-binding protein with curvature-generating and -sensing properties that participates in clathrin-dependent endocytosis of synaptic vesicle membranes. Endophilin also binds the GTPase dynamin and the phosphoinositide phosphatase synaptojanin and is thought to coordinate constriction of coated pits with membrane fission (via dynamin) and subsequent uncoating (via synaptojanin). We show that although synaptojanin is recruited by endophilin at bud necks before fission, the knockout of all three mouse endophilins results in the accumulation of clathrin-coated vesicles, but not of clathrin-coated pits, at synapses. The absence of endophilin impairs but does not abolish synaptic transmission and results in perinatal lethality, whereas partial endophilin absence causes severe neurological defects, including epilepsy and neurodegeneration. Our data support a model in which endophilin recruitment to coated pit necks, because of its curvature-sensing properties, primes vesicle buds for subsequent uncoating after membrane fission, without being critically required for the fission reaction itself.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures







Comment in
-
A surprisingly singular role for endophilin in synaptic vesicle recycling.Neuron. 2011 Nov 17;72(4):504-5. doi: 10.1016/j.neuron.2011.10.025. Neuron. 2011. PMID: 22099454
References
-
- Antonny B. Membrane deformation by protein coats. Curr Opin Cell Biol. 2006;18:386–394. - PubMed
-
- Blondeau F, Ritter B, Allaire PD, Wasiak S, Girard M, Hussain NK, Angers A, Legendre-Guillemin V, Roy L, Boismenu D, et al. Tandem MS analysis of brain clathrin-coated vesicles reveals their critical involvement in synaptic vesicle recycling. Proc Natl Acad Sci U S A. 2004;101:3833–3838. - PMC - PubMed
-
- Cremona O, Di Paolo G, Wenk MR, Luthi A, Kim WT, Takei K, Daniell L, Nemoto Y, Shears SB, Flavell RA, et al. Essential role of phosphoinositide metabolism in synaptic vesicle recycling. Cell. 1999;99:179–188. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
