

Viral microRNA targetome of KSHV-infected primary effusion lymphoma cell lines
- PMID: 22100165
- PMCID: PMC3222872
- DOI: 10.1016/j.chom.2011.09.012
Viral microRNA targetome of KSHV-infected primary effusion lymphoma cell lines
Abstract
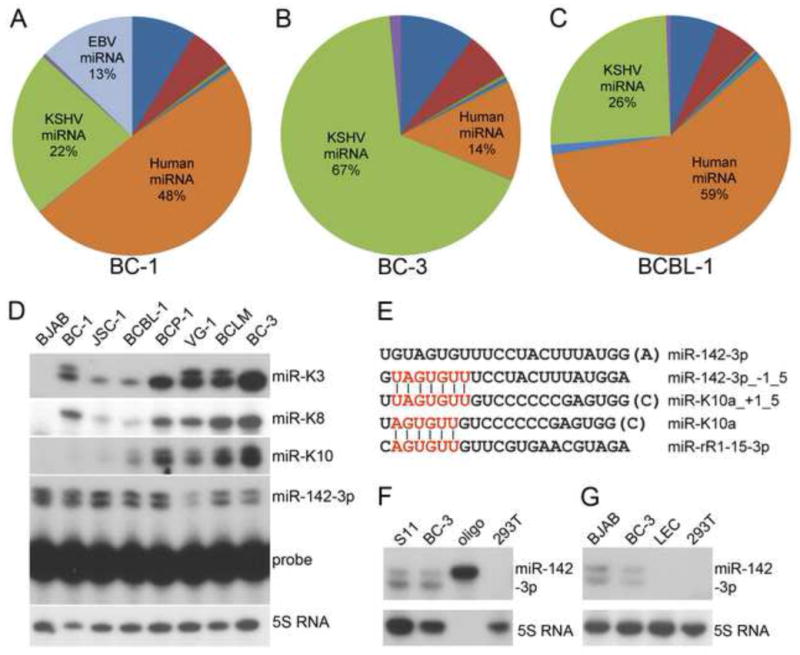
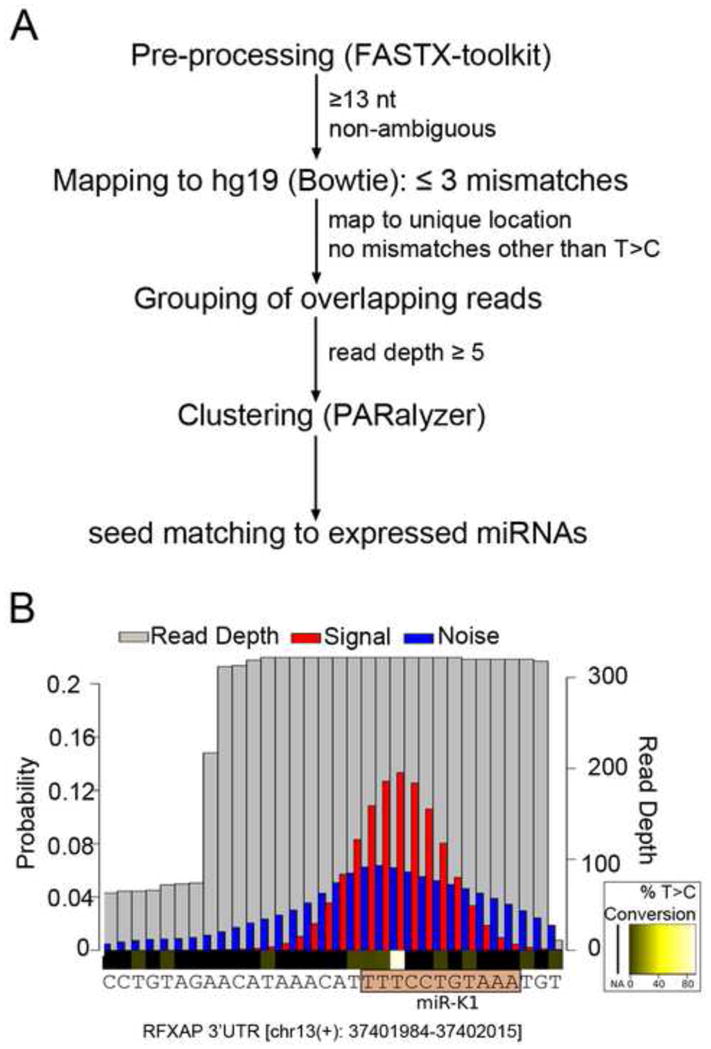
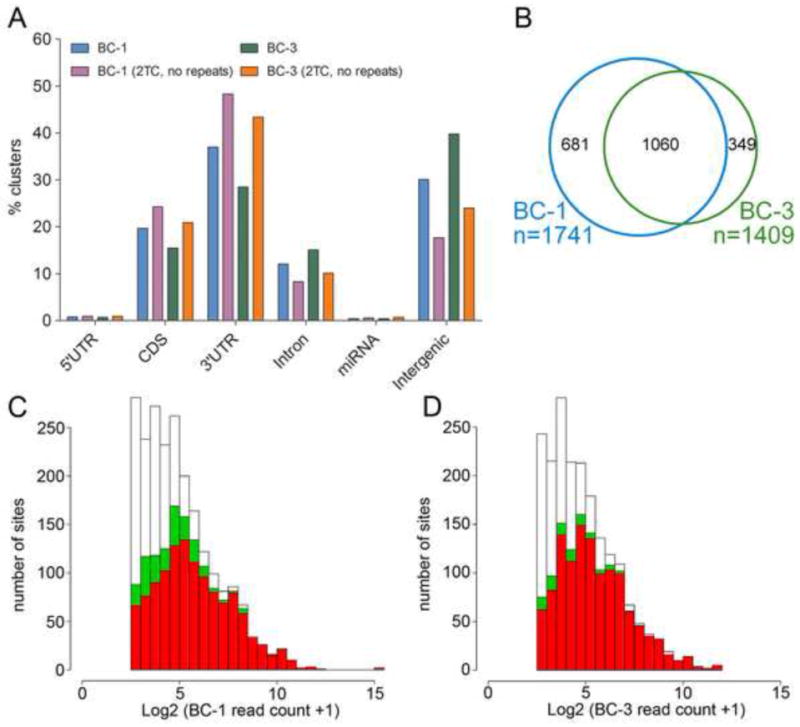
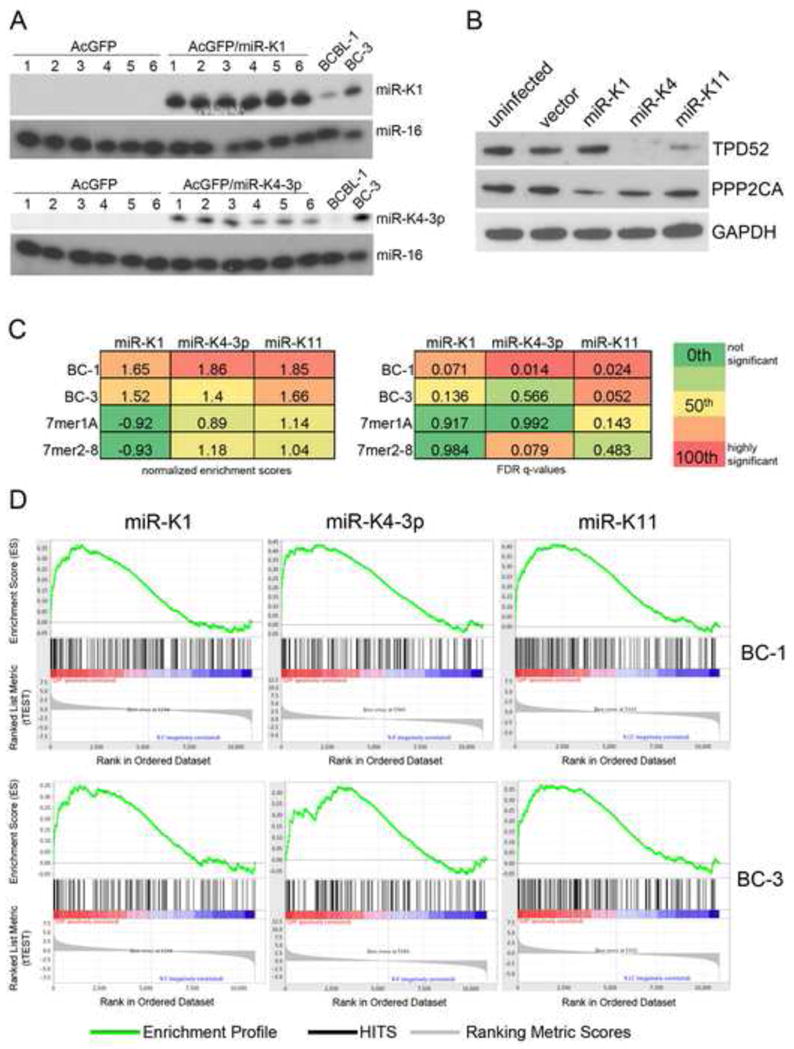
Primary effusion lymphoma (PEL) is caused by Kaposi's sarcoma-associated herpesvirus (KSHV) and frequently also harbors Epstein-Barr virus (EBV). The expression of KSHV- and EBV-encoded microRNAs (miRNAs) in PELs suggests a role for these miRNAs in latency and lymphomagenesis. Using PAR-CLIP, a technology which allows the direct and transcriptome-wide identification of miRNA targets, we delineate the target sites for all viral and cellular miRNAs expressed in PEL cell lines. The resulting data set revealed that KSHV miRNAs directly target more than 2000 cellular mRNAs, including many involved in pathways relevant to KSHV pathogenesis. Moreover, 58% of these mRNAs are also targeted by EBV miRNAs, via distinct binding sites. In addition to a known viral analog of cellular miR-155, we show that KSHV encodes a viral miRNA that mimics cellular miR-142-3p function. In summary, this study identifies an extensive list of KSHV miRNA targets, which are likely to influence viral replication and pathogenesis.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
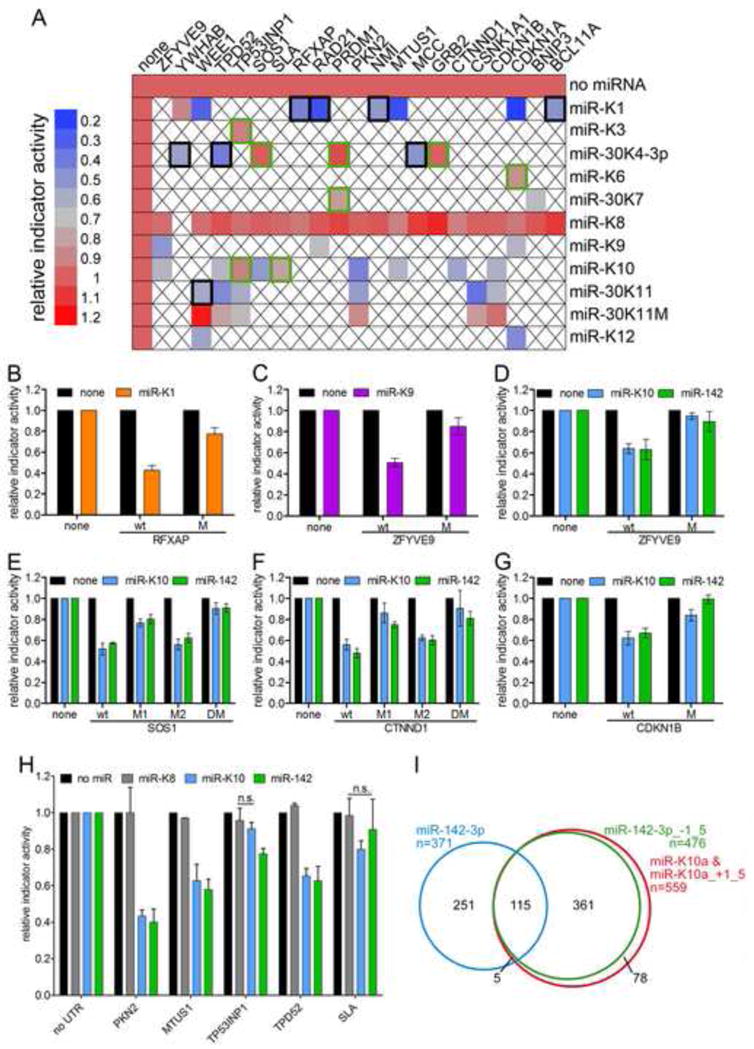
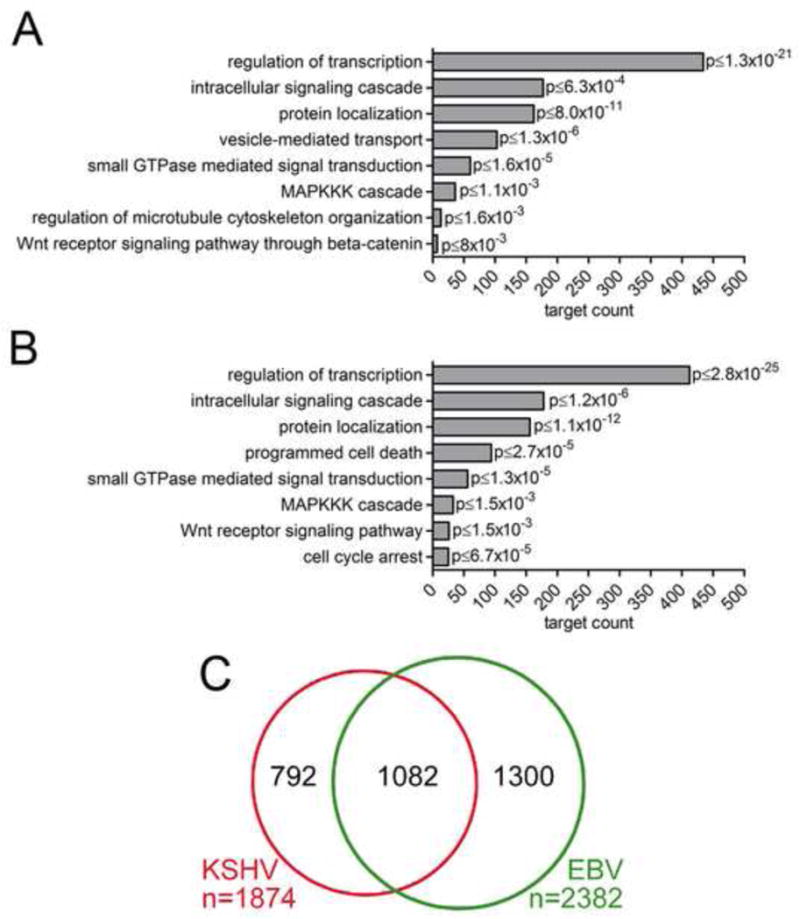
References
-
- Abend JR, Uldrick T, Ziegelbauer JM. Regulation of tumor necrosis factor-like weak inducer of apoptosis receptor protein (TWEAKR) expression by Kaposi’s sarcoma-associated herpesvirus microRNA prevents TWEAK-induced apoptosis and inflammatory cytokine expression. J Virol. 2010;84:12139–12151. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01-AI067968/AI/NIAID NIH HHS/United States
- RC1 CA145442/CA/NCI NIH HHS/United States
- RC1CA145442/CA/NCI NIH HHS/United States
- R00-CA-137860-02/CA/NCI NIH HHS/United States
- T32-CA009111/CA/NCI NIH HHS/United States
- R00 CA137860/CA/NCI NIH HHS/United States
- K99 CA137860/CA/NCI NIH HHS/United States
- GM073047/GM/NIGMS NIH HHS/United States
- P01 GM073047/GM/NIGMS NIH HHS/United States
- K99-CA-137860-01A1/CA/NCI NIH HHS/United States
- R01 AI067968/AI/NIAID NIH HHS/United States
- T32 CA009111/CA/NCI NIH HHS/United States
- MH08442/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
