

Cellular prion protein is essential for oligomeric amyloid-β-induced neuronal cell death
- PMID: 22100763
- PMCID: PMC3277312
- DOI: 10.1093/hmg/ddr542
Cellular prion protein is essential for oligomeric amyloid-β-induced neuronal cell death
Abstract
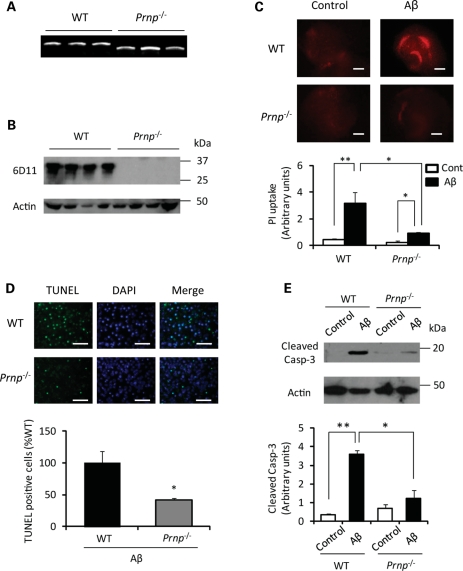
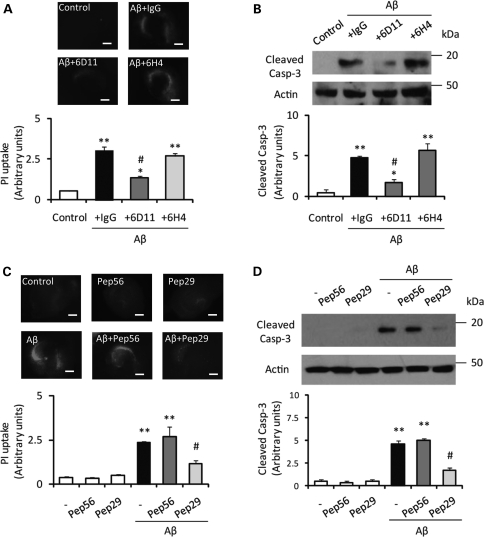
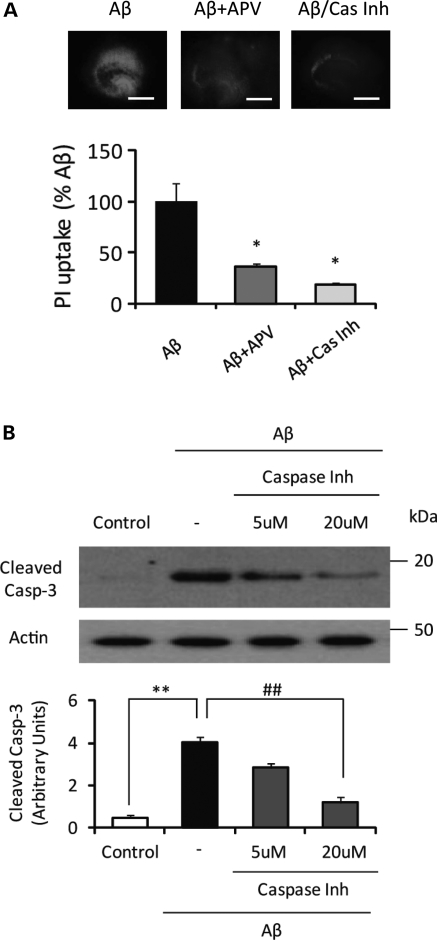
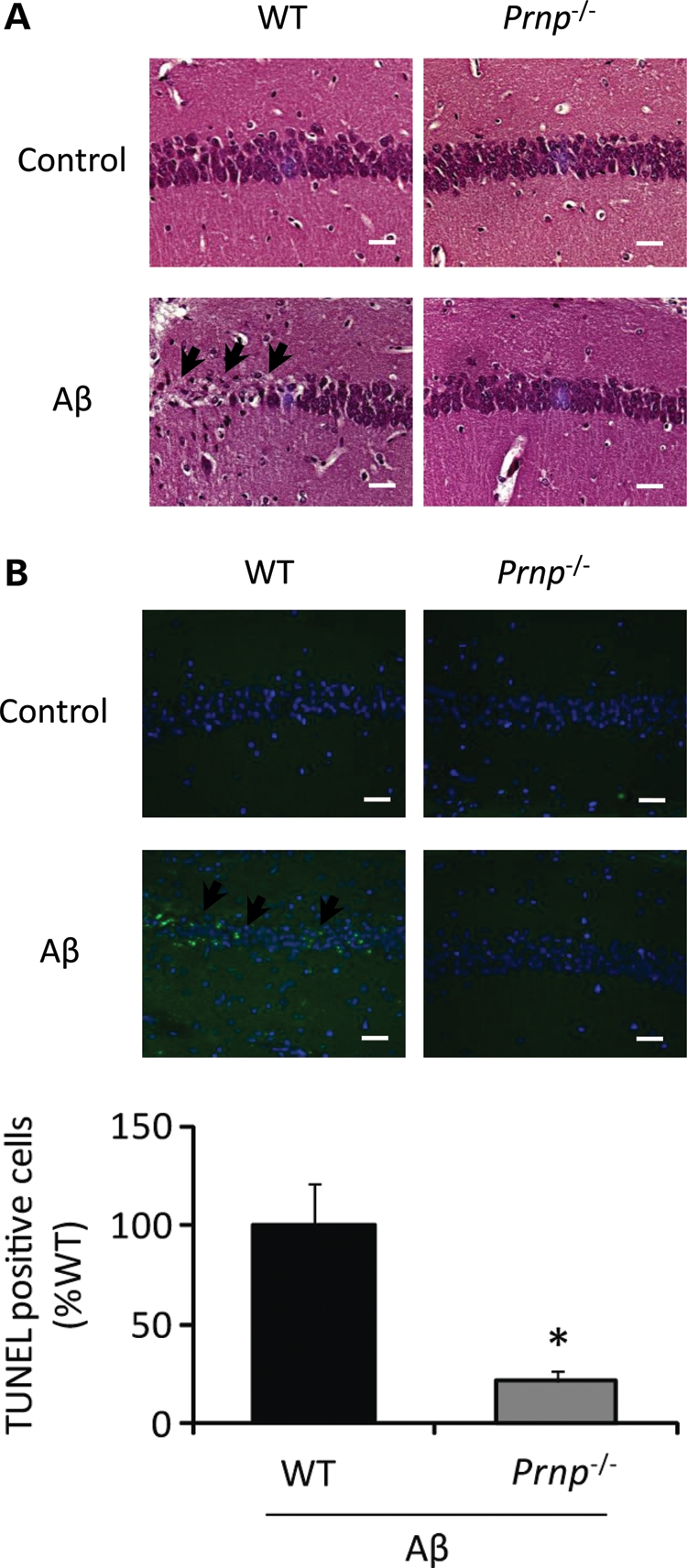
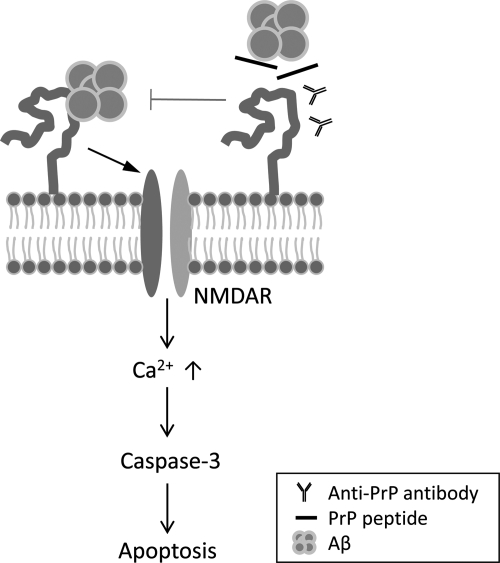
In Alzheimer disease (AD), amyloid-β (Aβ) oligomer is suggested to play a critical role in imitating neurodegeneration, although its pathogenic mechanism remains to be determined. Recently, the cellular prion protein (PrP(C)) has been reported to be an essential co-factor in mediating the neurotoxic effect of Aβ oligomer. However, these previous studies focused on the synaptic plasticity in either the presence or the absence of PrP(C) and no study to date has reported whether PrP(C) is required for the neuronal cell death, the most critical element of neurodegeneration in AD. Here, we show that Prnp(-/-) mice are resistant to the neurotoxic effect of Aβ oligomer in vivo and in vitro. Furthermore, application of an anti-PrP(C) antibody or PrP(C) peptide prevents Aβ oligomer-induced neurotoxicity. These findings are the first to demonstrate that PrP(C) is required for Aβ oligomer-induced neuronal cell death, the pathology essential to cognitive loss.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
