

Priming of adult pain responses by neonatal pain experience: maintenance by central neuroimmune activity
- PMID: 22102650
- PMCID: PMC3281475
- DOI: 10.1093/brain/awr288
Priming of adult pain responses by neonatal pain experience: maintenance by central neuroimmune activity
Abstract
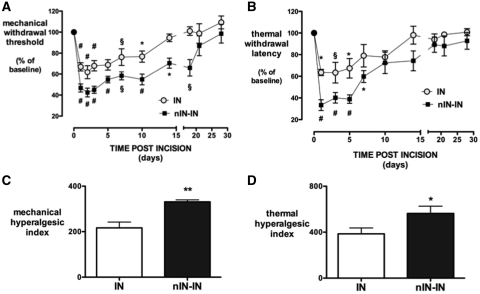
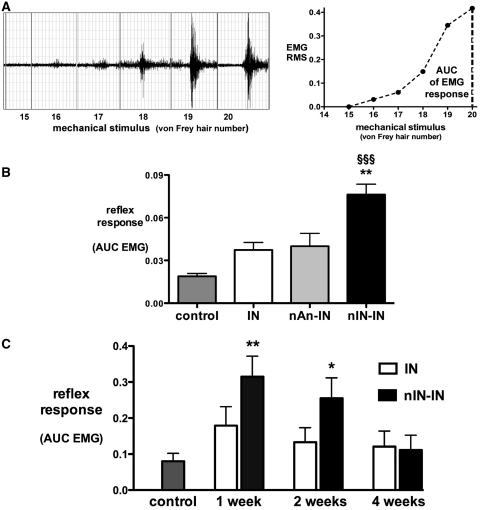
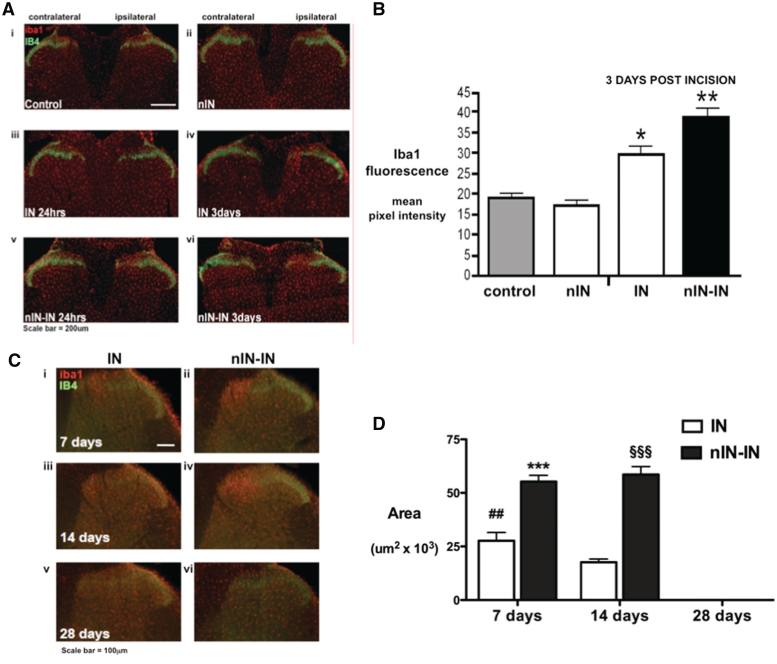
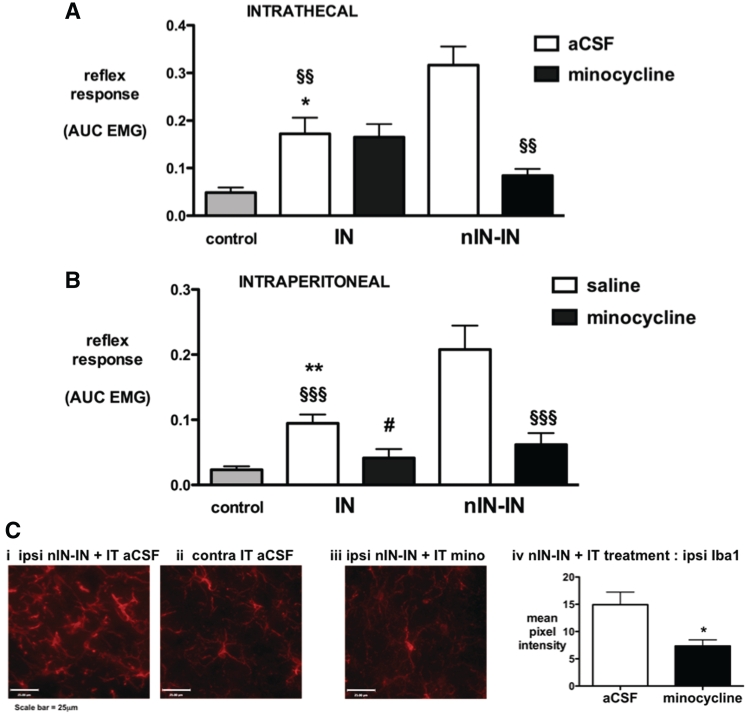
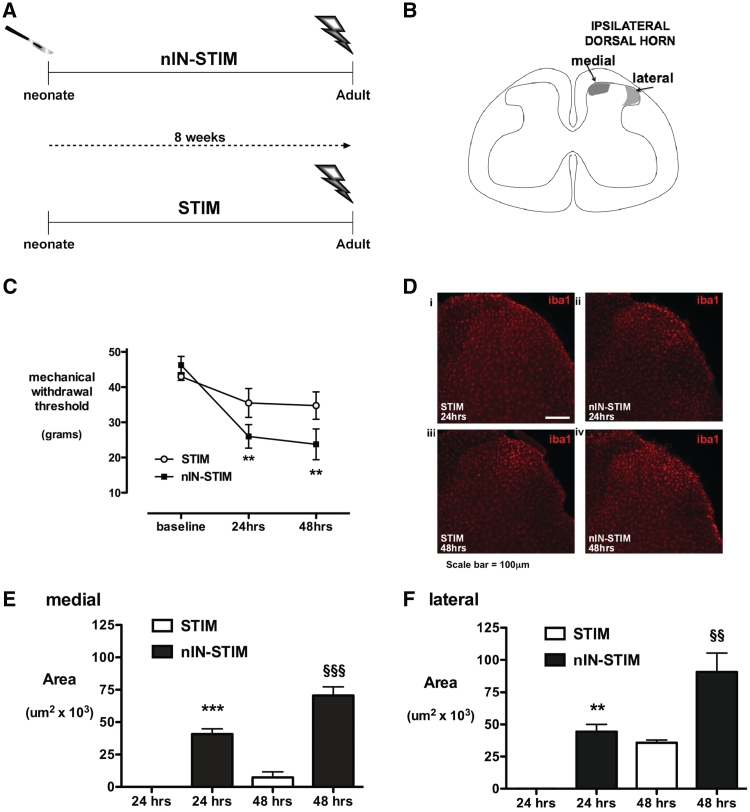
Adult brain connectivity is shaped by the balance of sensory inputs in early life. In the case of pain pathways, it is less clear whether nociceptive inputs in infancy can have a lasting influence upon central pain processing and adult pain sensitivity. Here, we show that adult pain responses in the rat are 'primed' by tissue injury in the neonatal period. Rats that experience hind-paw incision injury at 3 days of age, display an increased magnitude and duration of hyperalgesia following incision in adulthood when compared with those with no early life pain experience. This priming of spinal reflex sensitivity was measured by both reductions in behavioural withdrawal thresholds and increased flexor muscle electromyographic responses to graded suprathreshold hind-paw stimuli in the 4 weeks following adult incision. Prior neonatal injury also 'primed' the spinal microglial response to adult injury, resulting in an increased intensity, spatial distribution and duration of ionized calcium-binding adaptor molecule-1-positive microglial reactivity in the dorsal horn. Intrathecal minocycline at the time of adult injury selectively prevented both the hyperalgesia and early microglial reactivity associated with prior neonatal injury. The enhanced neuroimmune response seen in neonatally primed animals could also be demonstrated in the absence of peripheral tissue injury by direct electrical stimulation of tibial nerve fibres, confirming that centrally mediated mechanisms contribute to these long-term effects. These data suggest that early life injury may predispose individuals to enhanced sensitivity to painful events.
Figures
References
-
- Alexander GM, van Rijn MA, van Hilten JJ, Perreault MJ, Schwartzman RJ. Changes in cerebrospinal fluid levels of pro-inflammatory cytokines in CRPS. Pain. 2005;116:213–9. - PubMed
-
- Banik RK, Woo YC, Park SS, Brennan TJ. Strain and sex influence on pain sensitivity after plantar incision in the mouse. Anesthesiology. 2006;105:1246–53. - PubMed
-
- Bastos LF, Merlo LA, Rocha LT, Coelho MM. Characterization of the antinociceptive and anti-inflammatory activities of doxycycline and minocycline in different experimental models. Eur J Pharmacol. 2007;576:171–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
