

Early astrocytic atrophy in the entorhinal cortex of a triple transgenic animal model of Alzheimer's disease
- PMID: 22103264
- PMCID: PMC3243908
- DOI: 10.1042/AN20110025
Early astrocytic atrophy in the entorhinal cortex of a triple transgenic animal model of Alzheimer's disease
Abstract
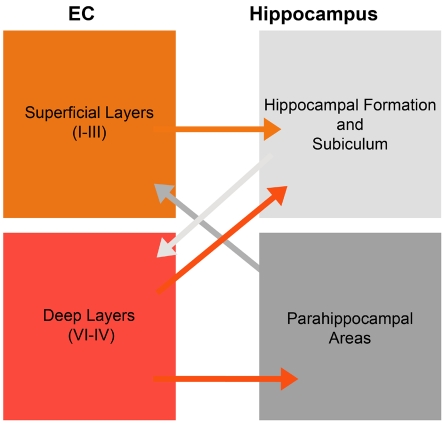
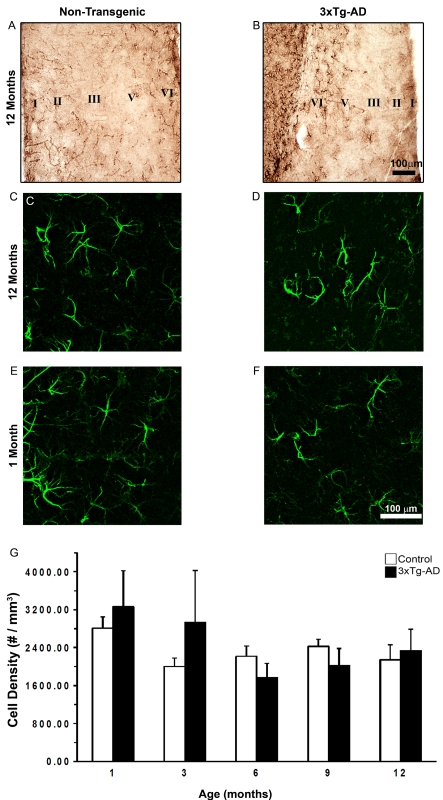
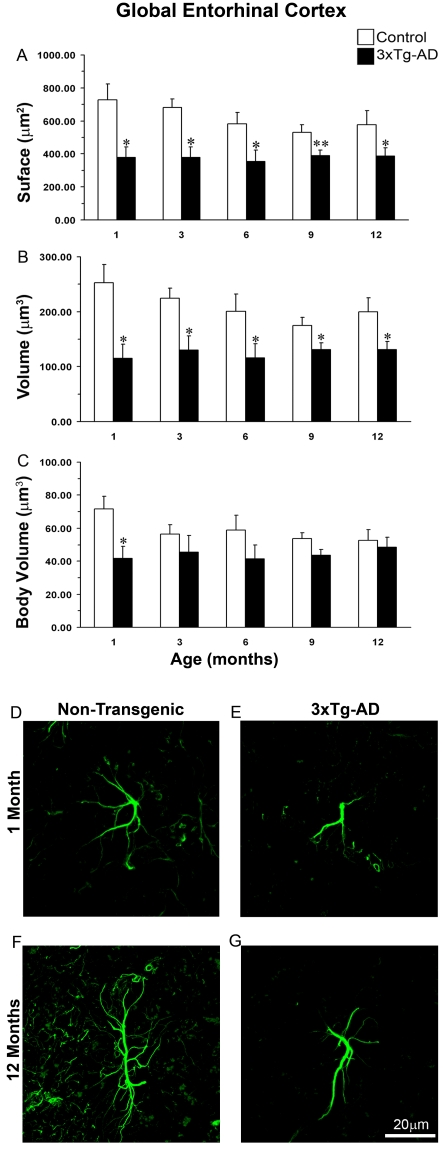
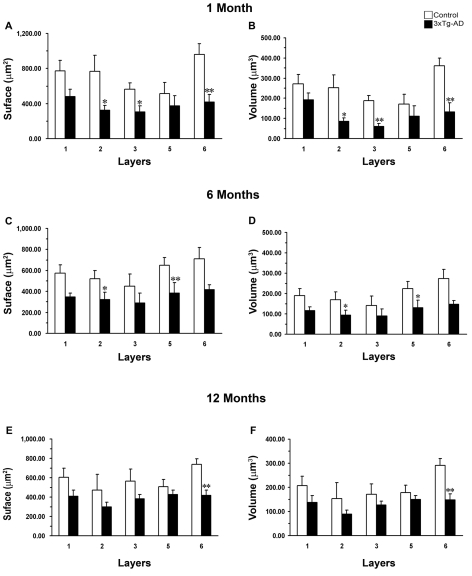
The EC (entorhinal cortex) is fundamental for cognitive and mnesic functions. Thus damage to this area appears as a key element in the progression of AD (Alzheimer's disease), resulting in memory deficits arising from neuronal and synaptic alterations as well as glial malfunction. In this paper, we have performed an in-depth analysis of astroglial morphology in the EC by measuring the surface and volume of the GFAP (glial fibrillary acidic protein) profiles in a triple transgenic mouse model of AD [3xTg-AD (triple transgenic mice of AD)]. We found significant reduction in both the surface and volume of GFAP-labelled profiles in 3xTg-AD animals from very early ages (1 month) when compared with non-Tg (non-transgenic) controls (48 and 54%, reduction respectively), which was sustained for up to 12 months (33 and 45% reduction respectively). The appearance of Aβ (amyloid β-peptide) depositions at 12 months of age did not trigger astroglial hypertrophy; nor did it result in the close association of astrocytes with senile plaques. Our results suggest that the AD progressive cognitive deterioration can be associated with an early reduction of astrocytic arborization and shrinkage of the astroglial domain, which may affect synaptic connectivity within the EC and between the EC and other brain regions. In addition, the EC seems to be particularly vulnerable to AD pathology because of the absence of evident astrogliosis in response to Aβ accumulation. Thus we can consider that targeting astroglial atrophy may represent a therapeutic strategy which might slow down the progression of AD.
Figures
References
-
- Araque A, Parpura V, Sanzgiri RP, Haydon PG. Tripartite synapses: glia, the unacknowledged partner. Trends Neurosci. 1999;22:208–215. - PubMed
-
- Bertoni-Freddari C, Sensi SL, Giogetti B, Balietti M, Di Stefano G, Canzoniero LM, Casoli T, Fattoretti P. Decreased presence of perforated synapses in a triple-transgenic mouse model of Alzheimer's disease. Rejuvenation Res. 2008;11:309–313. - PubMed
-
- Billings LM, Oddo S, Green KN, McGaugh JL, La Ferla FM. Intraneuronal Abeta causes the onset of early Alzheimer's disease-related cognitive deficits in transgenic mice. Neuron. 2005;45:675–688. - PubMed
-
- Braak H, Braak E. Neuropathological staging of Alzheimer-related changes. Acta Neuropathol. 1991;82:239–259. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
