

Persistent degenerative changes in thymic organ function revealed by an inducible model of organ regrowth
- PMID: 22103718
- PMCID: PMC4505612
- DOI: 10.1111/j.1474-9726.2011.00773.x
Persistent degenerative changes in thymic organ function revealed by an inducible model of organ regrowth
Abstract
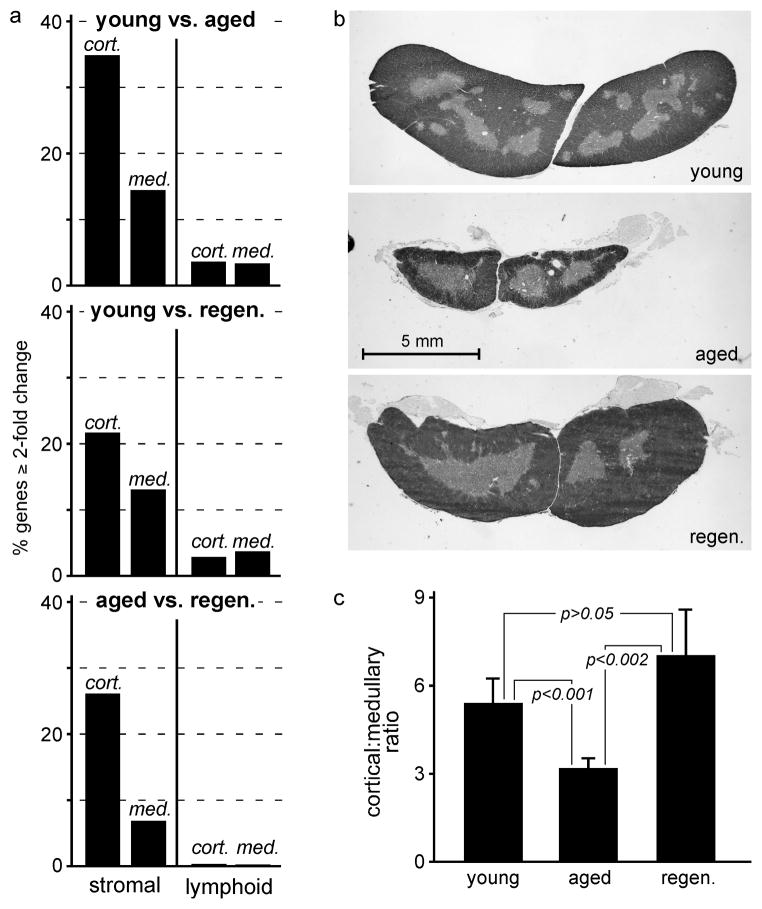
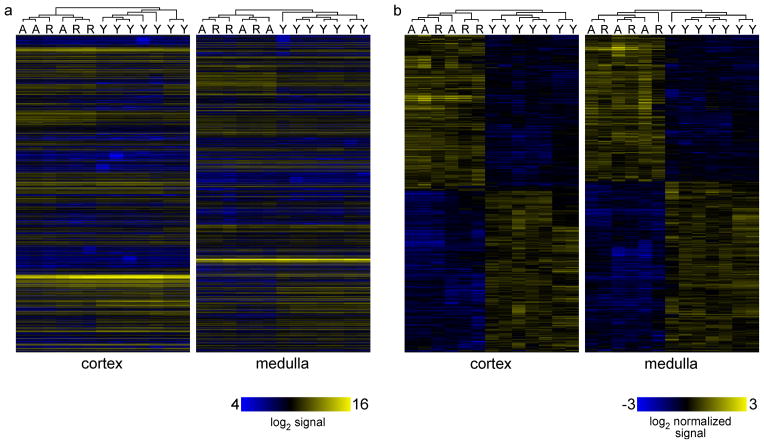
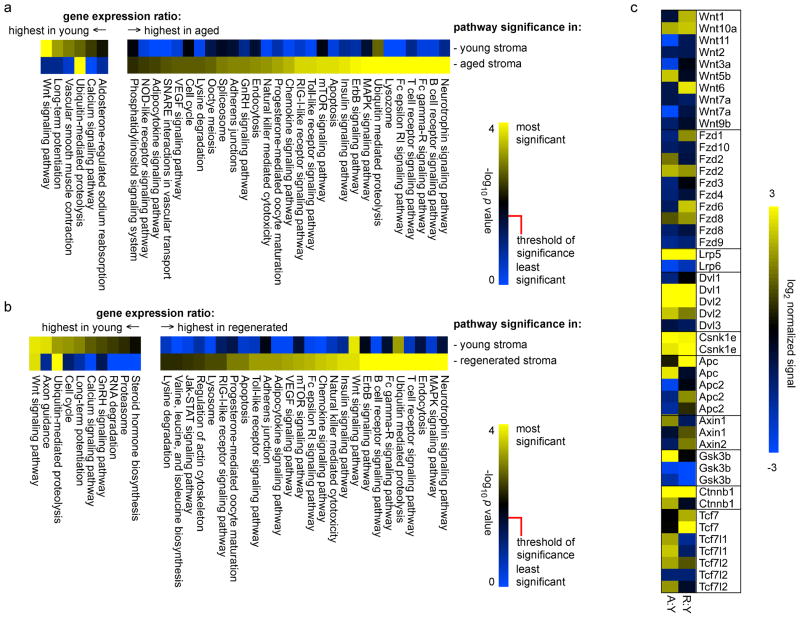
The thymus is the most rapidly aging tissue in the body, with progressive atrophy beginning as early as birth and not later than adolescence. Latent regenerative potential exists in the atrophic thymus, because certain stimuli can induce quantitative regrowth, but qualitative function of T lymphocytes produced by the regenerated organ has not been fully assessed. Using a genome-wide computational approach, we show that accelerated thymic aging is primarily a function of stromal cells, and that while overall cellularity of the thymus can be restored, many other aspects of thymic function cannot. Medullary islet complexity and tissue-restricted antigen expression decrease with age, representing potential mechanisms for age-related increases in autoimmune disease, but neither of these is restored by induced regrowth, suggesting that new T cells produced by the regrown thymus will probably include more autoreactive cells. Global analysis of stromal gene expression profiles implicates widespread changes in Wnt signaling as the most significant hallmark of degeneration, changes that once again persist even at peak regrowth. Consistent with the permanent nature of age-related molecular changes in stromal cells, induced thymic regrowth is not durable, with the regrown organ returning to an atrophic state within 2 weeks of reaching peak size. Our findings indicate that while quantitative regrowth of the thymus is achievable, the changes associated with aging persist, including potential negative implications for autoimmunity.
© 2011 The Authors. Aging Cell © 2011 Blackwell Publishing Ltd/Anatomical Society of Great Britain and Ireland.
Figures


References
-
- Balciunaite G, Keller MP, Balciunaite E, Piali L, Zuklys S, Mathieu YD, Gill J, Boyd R, Sussman DJ, Hollander GA. Wnt glycoproteins regulate the expression of FoxN1, the gene defective in nude mice. Nat Immunol. 2002;3:1102–1108. - PubMed
-
- Besedovsky HO, Sorkin E. Thymus involvement in female sexual maturation. Nature. 1974;249:356–358. - PubMed
-
- Boehm T, Bleul CC. The evolutionary history of lymphoid organs. Nat Immunol. 2007;8:131–135. - PubMed
-
- Brelinska R, Malendowicz LK, Malinska A, Kowalska K. Characteristics of age-related changes in rat thymus: morphometric analysis and epithelial cell network in various thymic compartments. Biogerontology. 2008;9:93–108. - PubMed
-
- Brunelli R, Frasca D, Spano M, Zichella L, Doria G. Gonadectomy in old mice induces thymus regeneration but does not recover mitotic responsiveness. Ann N Y Acad Sci. 1992;673:252–255. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
