

A phylogeny and molecular barcodes for Caenorhabditis, with numerous new species from rotting fruits
- PMID: 22103856
- PMCID: PMC3277298
- DOI: 10.1186/1471-2148-11-339
A phylogeny and molecular barcodes for Caenorhabditis, with numerous new species from rotting fruits
Abstract
Background: The nematode Caenorhabditis elegans is a major laboratory model in biology. Only ten Caenorhabditis species were available in culture at the onset of this study. Many of them, like C. elegans, were mostly isolated from artificial compost heaps, and their more natural habitat was unknown.
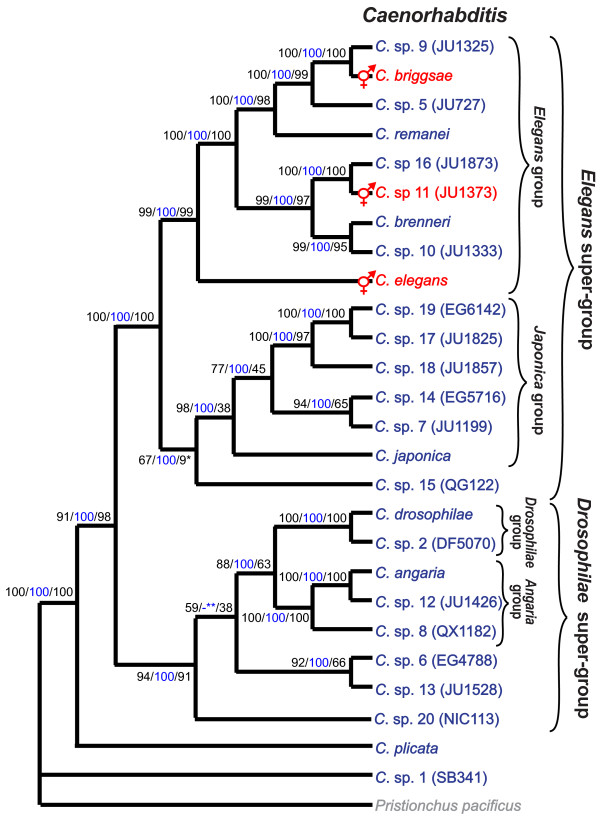
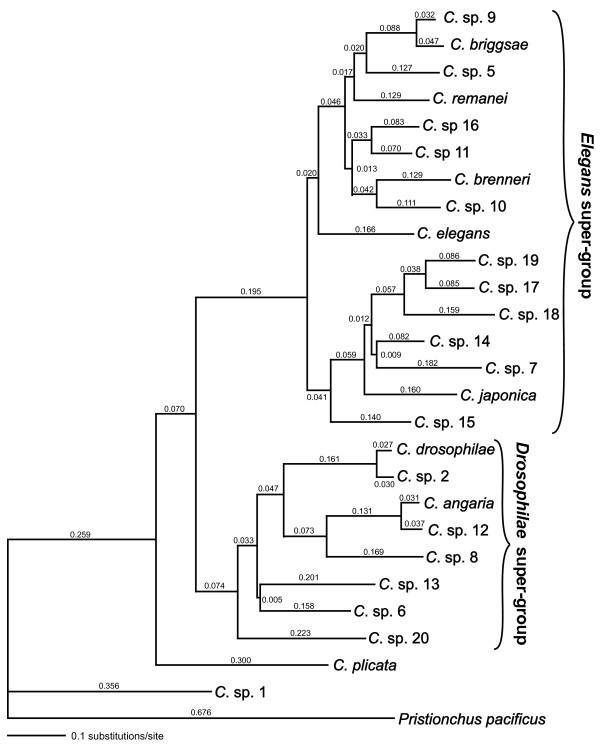
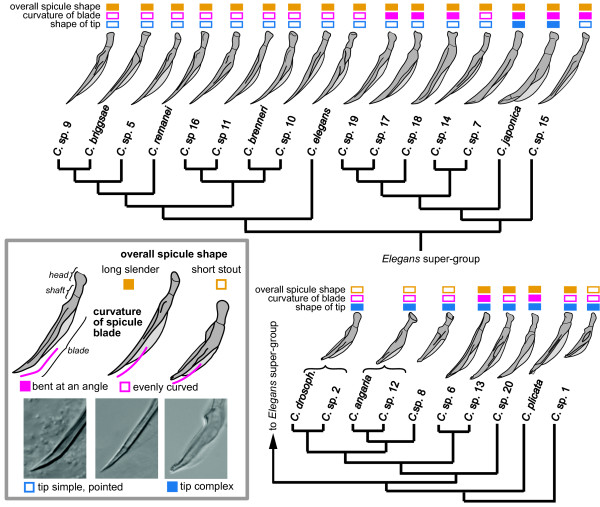
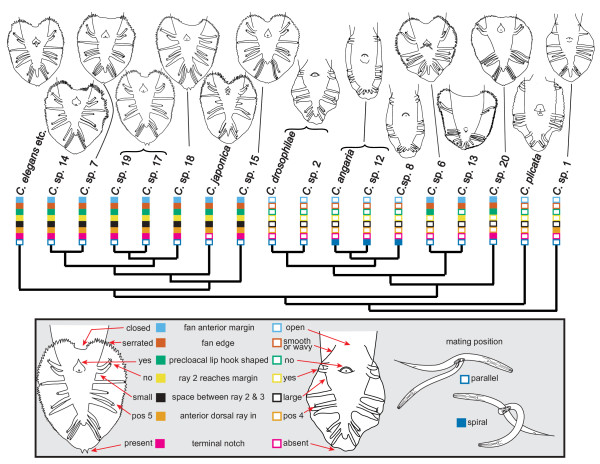
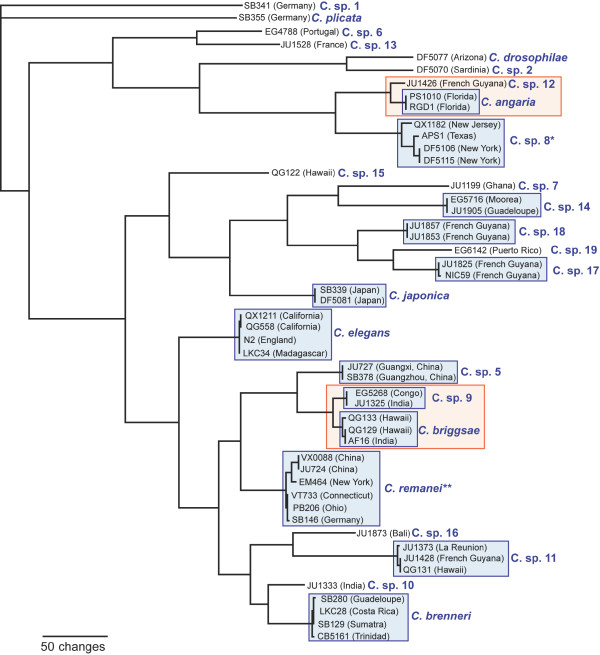
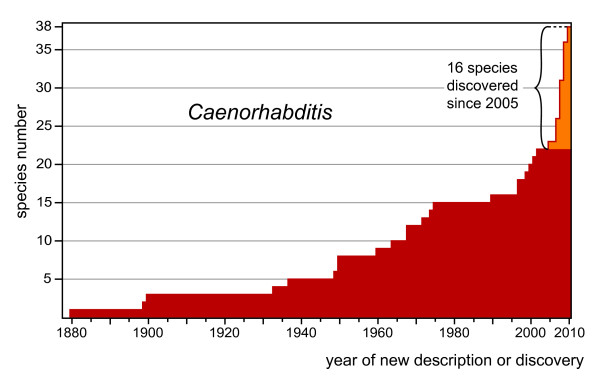
Results: Caenorhabditis nematodes were found to be proliferating in rotten fruits, flowers and stems. By collecting a large worldwide set of such samples, 16 new Caenorhabditis species were discovered. We performed mating tests to establish biological species status and found some instances of semi-fertile or sterile hybrid progeny. We established barcodes for all species using ITS2 rDNA sequences. By obtaining sequence data for two rRNA and nine protein-coding genes, we determined the likely phylogenetic relationships among the 26 species in culture. The new species are part of two well-resolved sister clades that we call the Elegans super-group and the Drosophilae super-group. We further scored phenotypic characters such as reproductive mode, mating behavior and male tail morphology, and discuss their congruence with the phylogeny. A small space between rays 2 and 3 evolved once in the stem species of the Elegans super-group; a narrow fan and spiral copulation evolved once in the stem species of C. angaria, C. sp. 8 and C. sp. 12. Several other character changes occurred convergently. For example, hermaphroditism evolved three times independently in C. elegans, C. briggsae and C. sp. 11. Several species can co-occur in the same location or even the same fruit. At the global level, some species have a cosmopolitan distribution: C. briggsae is particularly widespread, while C. elegans and C. remanei are found mostly or exclusively in temperate regions, and C. brenneri and C. sp. 11 exclusively in tropical zones. Other species have limited distributions, for example C. sp. 5 appears to be restricted to China, C. sp. 7 to West Africa and C. sp. 8 to the Eastern United States.
Conclusions: Caenorhabditis are "fruit worms", not soil nematodes. The 16 new species provide a resource and their phylogeny offers a framework for further studies into the evolution of genomic and phenotypic characters.
Figures


References
-
- Barrière A, Félix M-A. Natural variation and population genetics of C. elegans. The C. elegans Research Community. January 9, 2006. http://www.wormbook.org/ - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
