

Toxoplasma gondii triggers release of human and mouse neutrophil extracellular traps
- PMID: 22104111
- PMCID: PMC3264325
- DOI: 10.1128/IAI.05730-11
Toxoplasma gondii triggers release of human and mouse neutrophil extracellular traps
Abstract
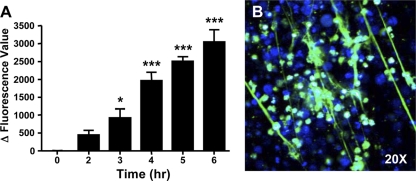
Neutrophils have recently been shown to release DNA-based extracellular traps that contribute to microbicidal killing and have also been implicated in autoimmunity. The role of neutrophil extracellular trap (NET) formation in the host response to nonbacterial pathogens has received much less attention. Here, we show that the protozoan pathogen Toxoplasma gondii elicits the production of NETs from human and mouse neutrophils. Tachyzoites of each of the three major parasite strain types were efficiently entrapped within NETs, resulting in decreased parasite viability. We also show that Toxoplasma activates a MEK-extracellular signal-regulated kinase (ERK) pathway in neutrophils and that the inhibition of this pathway leads to decreased NET formation. To determine if Toxoplasma induced NET formation in vivo, we employed a mouse intranasal infection model. We found that the administration of tachyzoites by this route induced a rapid tissue recruitment of neutrophils with evidence of extracellular DNA release. Taken together, these data indicate a role for NETs in the host innate response to protozoan infection. We propose that NET formation limits infection by direct microbicidal effects on Toxoplasma as well as by interfering with the ability of the parasite to invade target host cells.
Figures









References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
