

Intra-membrane oligomerization and extra-membrane oligomerization of amyloid-β peptide are competing processes as a result of distinct patterns of motif interplay
- PMID: 22105077
- PMCID: PMC3249129
- DOI: 10.1074/jbc.M111.281295
Intra-membrane oligomerization and extra-membrane oligomerization of amyloid-β peptide are competing processes as a result of distinct patterns of motif interplay
Abstract
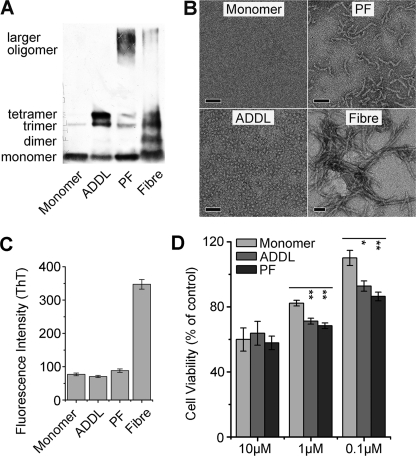
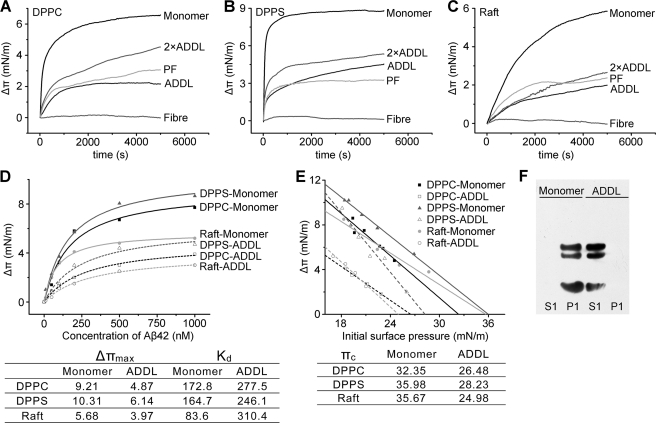
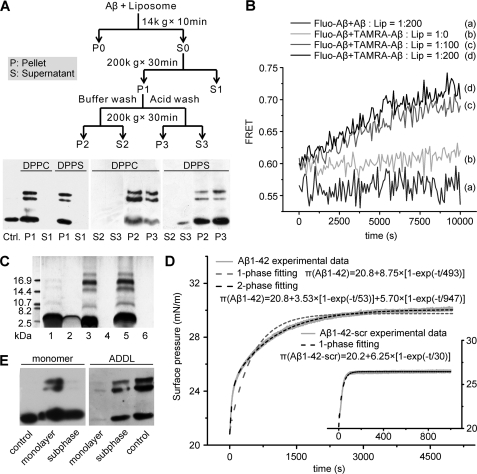
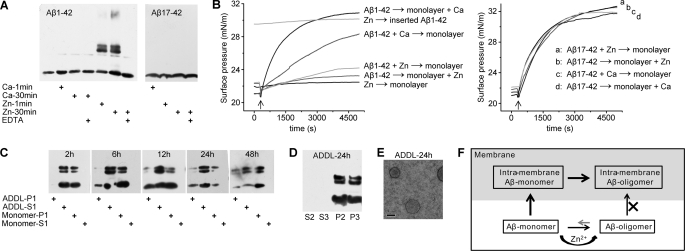
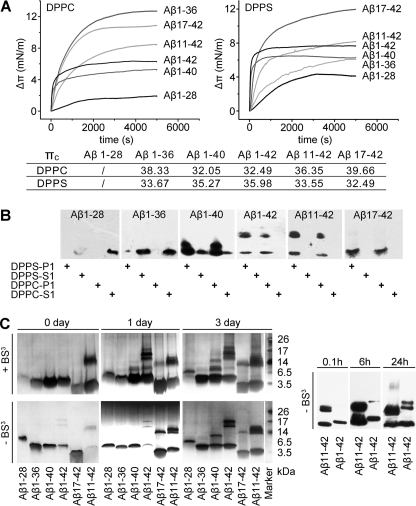
Soluble oligomers of amyloid-β peptide (Aβ) are emerging as the primary neurotoxic species in Alzheimer disease, however, whether the membrane is among their direct targets that mediate the downstream adverse effects remains elusive. Herein, we show that multiple soluble oligomeric Aβ preparations, including Aβ-derived diffusible ligand, protofibril, and zinc-induced Aβ oligomer, exhibit much weaker capability to insert into the membrane than Aβ monomer. Aβ monomers prefer incorporating into membrane rather than oligomerizing in solution, and such preference can be reversed by the aggregation-boosting factor, zinc ion. Further analyses indicate that the membrane-embedded oligomers of Aβ are derived from rapid assembly of inserted monomers but not due to the insertion of soluble Aβ oligomers. By comparing the behavior of a panel of Aβ truncation variants, we demonstrate that the intra- and extra-membrane oligomerization are mutually exclusive processes that proceed through distinct motif interplay, both of which require the action of amino acids 37-40/42 to overcome the auto-inhibitory interaction between amino acids 29-36 and the N-terminal portion albeit via different mechanisms. These results indicate that intra- and extra-membrane oligomerization of Aβ are competing processes and emphasize a critical regulation of membrane on the behavior of Aβ monomer and soluble oligomers, which may determine distinct neurotoxic mechanisms.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
