

Affinity maturation of antibodies requires integrity of the adult thymus
- PMID: 22105515
- PMCID: PMC4855521
- DOI: 10.1002/eji.201141889
Affinity maturation of antibodies requires integrity of the adult thymus
Abstract
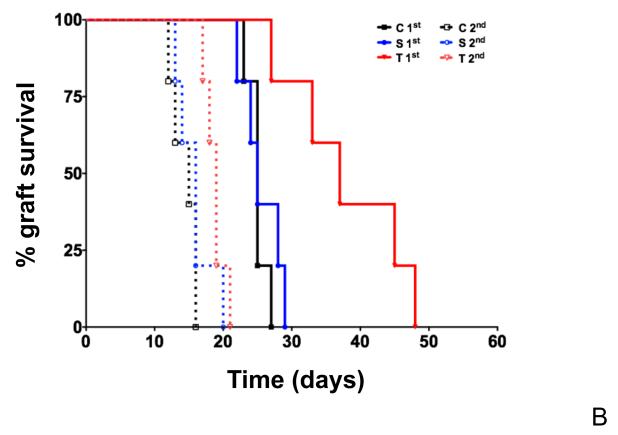
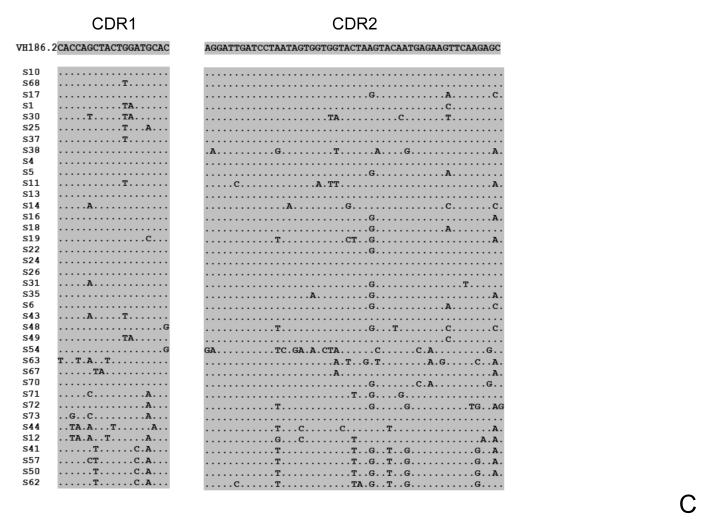
The generation of B-cell responses to proteins requires a functional thymus to produce CD4(+) T cells which helps in the activation and differentiation of B cells. Because the mature T-cell repertoire has abundant cells with the helper phenotype, one might predict that in mature individuals, the generation of B-cell memory would proceed independently of the thymus. Contrary to that prediction, we show here that the removal of the thymus after the establishment of the T-cell compartment or sham surgery without removal of the thymus impairs the affinity maturation of antibodies. Because removal or manipulation of the thymus did not decrease the frequency of mutation of the Ig variable heavy chain exons encoding antigen-specific antibodies, we conclude that the thymus controls affinity maturation of antibodies in the mature individual by facilitating the selection of B cells with high-affinity antibodies.
Copyright © 2012 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Figures











References
-
- Manz RA, Arce S, Cassese G, Hauser AE, Hiepe F, Radbruch A. Humoral immunity and long-lived plasma cells. Curr Opin Immunol. 2002;14:517–521. - PubMed
-
- Miller JF. Immunologic function of the thymus. Lancet. 1961;2:748–749. - PubMed
-
- Miller JF, De Burgh PM, Grant GA. Thymus and the production of antibody-plaque-forming cells. Nature. 1965;208:1332–1334. - PubMed
-
- Miller JF. Effect of thymectomy in adult mice on immunological responsiveness. Nature. 1965;208:1337–1338. - PubMed
-
- Metcalf D. Delayed effect of thymectomy in adult life on immunological competence. Nature. 1965;208:1336. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
