

Arenavirus Z protein controls viral RNA synthesis by locking a polymerase-promoter complex
- PMID: 22106304
- PMCID: PMC3241799
- DOI: 10.1073/pnas.1112742108
Arenavirus Z protein controls viral RNA synthesis by locking a polymerase-promoter complex
Abstract
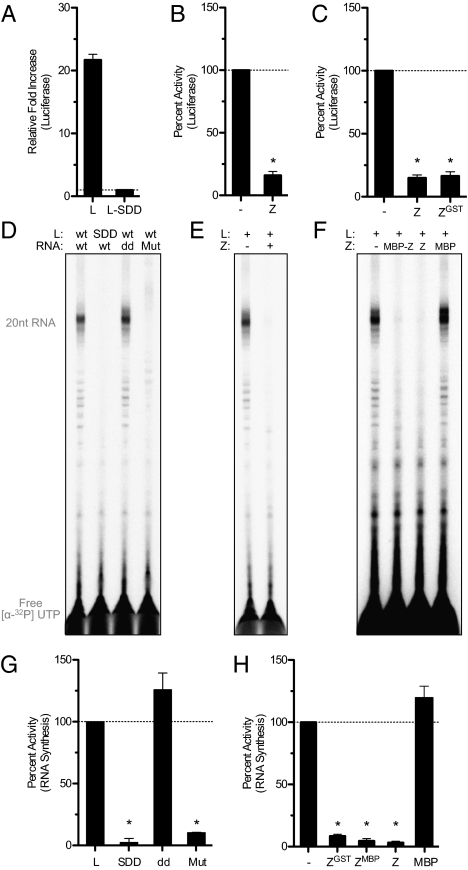
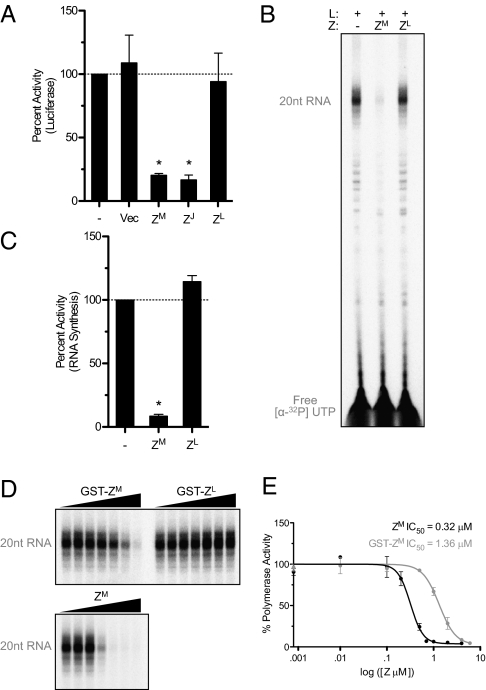
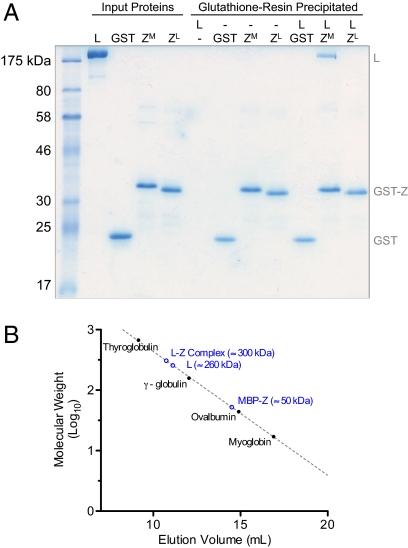
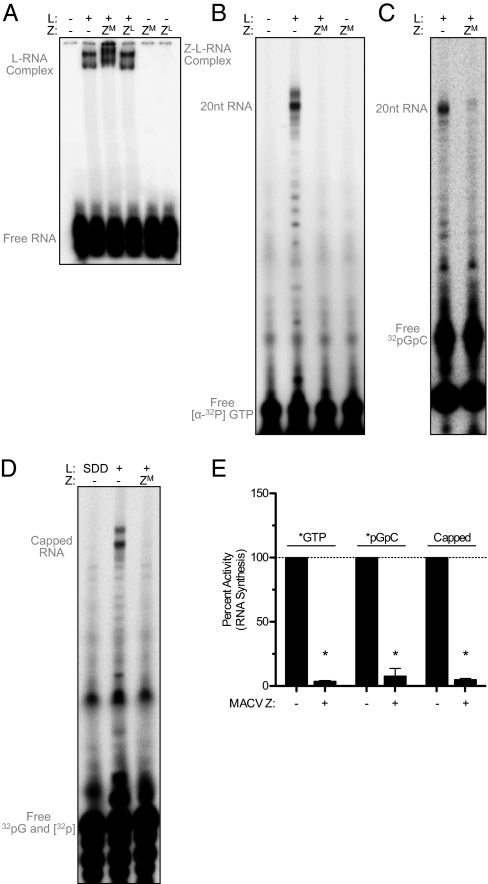
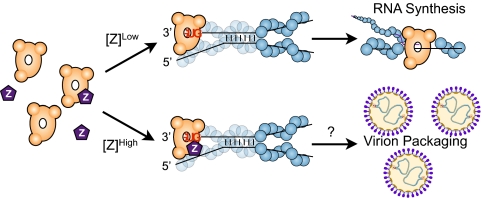
Arenaviruses form a noncytolytic infection in their rodent hosts, yet can elicit severe hemorrhagic disease in humans. How arenaviruses regulate gene expression remains unclear, and further understanding may provide insight into the dichotomy of these disparate infection processes. Here we reconstitute arenavirus RNA synthesis initiation and gene expression regulation in vitro using purified components and demonstrate a direct role of the viral Z protein in controlling RNA synthesis. Our data reveal that Z forms a species-specific complex with the viral polymerase (L) and inhibits RNA synthesis initiation by impairing L catalytic activity. This Z-L complex locks the viral polymerase in a promoter-bound, catalytically inactive state and may additionally ensure polymerase packaging during virion maturation. Z modulates host factors involved in cellular translation, proliferation, and antiviral signaling. Our data defines an additional role in governing viral RNA synthesis, revealing Z as the center of a network of host and viral connections that regulates viral gene expression.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Tjian R. The biochemistry of transcription in eukaryotes: A paradigm for multisubunit regulatory complexes. Philos Trans R Soc Lond B Biol Sci. 1996;351:491–499. - PubMed
-
- Kumar A, Patel SS. Inhibition of T7 RNA polymerase: Transcription initiation and transition from initiation to elongation are inhibited by T7 lysozyme via a ternary complex with RNA polymerase and promoter DNA. Biochemistry. 1997;36:13954–13962. - PubMed
-
- Zhang X, Studier FW. Mechanism of inhibition of bacteriophage T7 RNA polymerase by T7 lysozyme. J Mol Biol. 1997;269:10–27. - PubMed
-
- Childs JE, Peters CJ. Ecology and epidemiology of arenaviruses and their hosts. In: Salvato MS, editor. The Arenaviridae. New York: Plenum; 1993. pp. 331–373.
-
- Geisbert TW, Jahrling PB. Exotic emerging viral diseases: Progress and challenges. Nat Med. 2004;10(12, Suppl):S110–S121. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
