

BLM helicase facilitates RNA polymerase I-mediated ribosomal RNA transcription
- PMID: 22106380
- PMCID: PMC3277314
- DOI: 10.1093/hmg/ddr545
BLM helicase facilitates RNA polymerase I-mediated ribosomal RNA transcription
Abstract
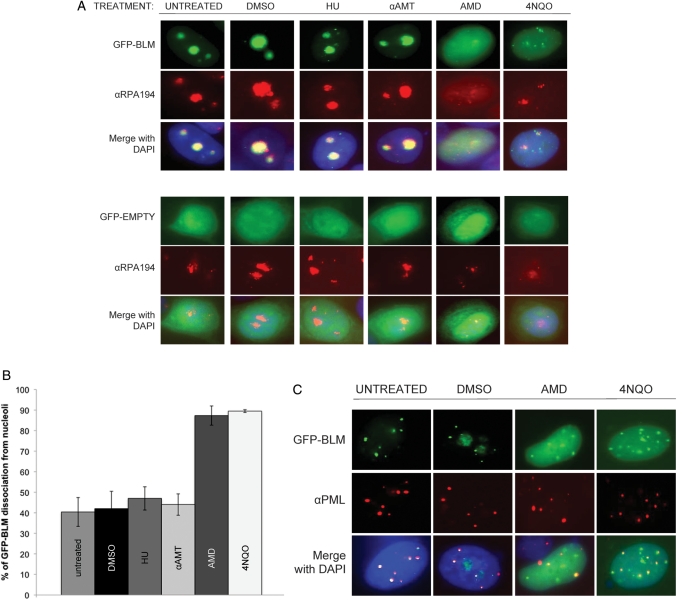
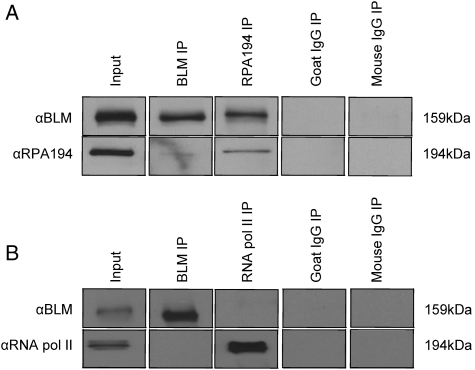
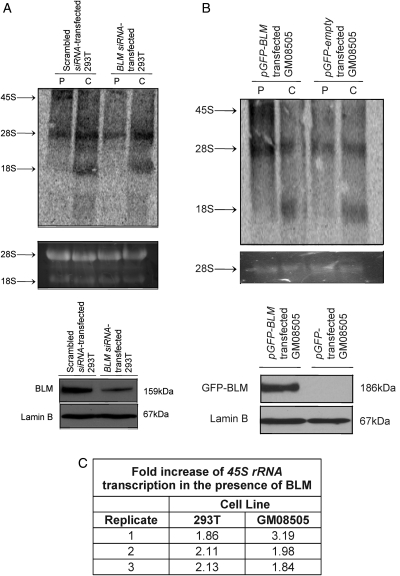
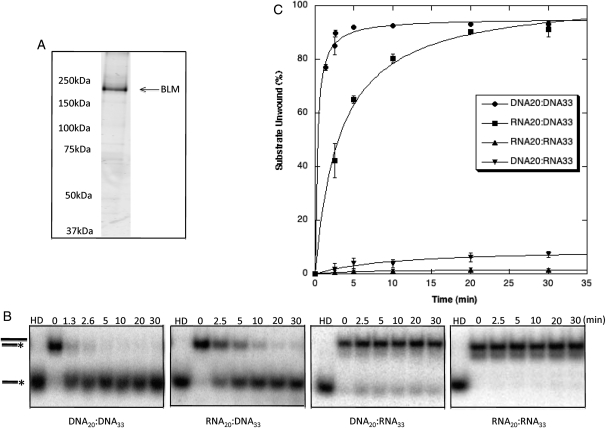
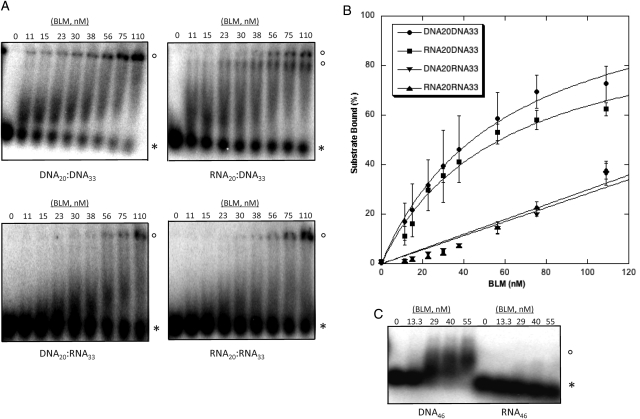
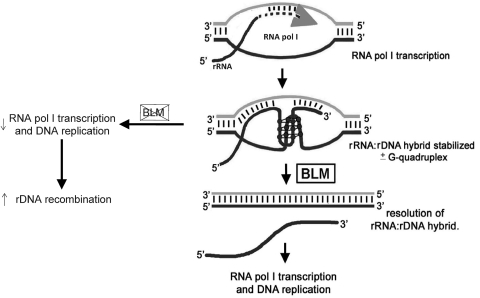
Bloom's syndrome (BS) is an autosomal recessive disorder that is invariably characterized by severe growth retardation and cancer predisposition. The Bloom's syndrome helicase (BLM), mutations of which lead to BS, localizes to promyelocytic leukemia protein bodies and to the nucleolus of the cell, the site of RNA polymerase I-mediated ribosomal RNA (rRNA) transcription. rRNA transcription is fundamental for ribosome biogenesis and therefore protein synthesis, cellular growth and proliferation; its inhibition limits cellular growth and proliferation as well as bodily growth. We report that nucleolar BLM facilitates RNA polymerase I-mediated rRNA transcription. Immunofluorescence studies demonstrate the dependance of BLM nucleolar localization upon ongoing RNA polymerase I-mediated rRNA transcription. In vivo protein co-immunoprecipitation demonstrates that BLM interacts with RPA194, a subunit of RNA polymerase I. (3)H-uridine pulse-chase assays demonstrate that BLM expression is required for efficient rRNA transcription. In vitro helicase assays demonstrate that BLM unwinds GC-rich rDNA-like substrates that form in the nucleolus and normally inhibit progression of the RNA polymerase I transcription complex. These studies suggest that nucleolar BLM modulates rDNA structures in association with RNA polymerase I to facilitate RNA polymerase I-mediated rRNA transcription. Given the intricate relationship between rDNA metabolism and growth, our data may help in understanding the etiology of proportional dwarfism in BS.
Figures
Similar articles
-
Collaborating functions of BLM and DNA topoisomerase I in regulating human rDNA transcription.Mutat Res. 2013 Mar-Apr;743-744:89-96. doi: 10.1016/j.mrfmmm.2012.12.002. Epub 2012 Dec 19. Mutat Res. 2013. PMID: 23261817 Free PMC article.
-
WRN helicase accelerates the transcription of ribosomal RNA as a component of an RNA polymerase I-associated complex.Oncogene. 2002 Apr 11;21(16):2447-54. doi: 10.1038/sj.onc.1205334. Oncogene. 2002. PMID: 11971179
-
HERC2 inactivation abrogates nucleolar localization of RecQ helicases BLM and WRN.Sci Rep. 2021 Jan 11;11(1):360. doi: 10.1038/s41598-020-79715-y. Sci Rep. 2021. PMID: 33432007 Free PMC article.
-
Beyond rRNA: nucleolar transcription generates a complex network of RNAs with multiple roles in maintaining cellular homeostasis.Genes Dev. 2022 Aug 1;36(15-16):876-886. doi: 10.1101/gad.349969.122. Genes Dev. 2022. PMID: 36207140 Free PMC article. Review.
-
The nucleolus—guardian of cellular homeostasis and genome integrity.Chromosoma. 2013 Dec;122(6):487-97. doi: 10.1007/s00412-013-0430-0. Chromosoma. 2013. PMID: 24022641 Review.
Cited by
-
The BLM helicase is a new therapeutic target in multiple myeloma involved in replication stress survival and drug resistance.Front Immunol. 2022 Dec 9;13:983181. doi: 10.3389/fimmu.2022.983181. eCollection 2022. Front Immunol. 2022. PMID: 36569948 Free PMC article.
-
Dynamic nucleoplasmic and nucleolar localization of mammalian RNase H1 in response to RNAP I transcriptional R-loops.Nucleic Acids Res. 2017 Oct 13;45(18):10672-10692. doi: 10.1093/nar/gkx710. Nucleic Acids Res. 2017. PMID: 28977560 Free PMC article.
-
An overview of RecQ helicases and related diseases.Aging (Albany NY). 2025 Jul 25;17(7):1881-1907. doi: 10.18632/aging.206291. Epub 2025 Jul 25. Aging (Albany NY). 2025. PMID: 40728512 Free PMC article. Review.
-
RECQ5 mediates pre-rRNA processing in nucleolus.Nucleic Acids Res. 2025 Aug 11;53(15):gkaf766. doi: 10.1093/nar/gkaf766. Nucleic Acids Res. 2025. PMID: 40823811 Free PMC article.
-
Collaborating functions of BLM and DNA topoisomerase I in regulating human rDNA transcription.Mutat Res. 2013 Mar-Apr;743-744:89-96. doi: 10.1016/j.mrfmmm.2012.12.002. Epub 2012 Dec 19. Mutat Res. 2013. PMID: 23261817 Free PMC article.
References
-
- Schwarzacher H.G., Wachtler F. The nucleolus. Anat. Embryol. 1993;188:515–536. - PubMed
-
- Ayrault O., Andrique L., Fauvin D., Eymin B., Gazzeri S., Seite P. Human tumor suppressor p14ARF negatively regulates rRNA transcription and inhibits UBF1 transcription factor phosphorylation. Oncogene. 2006;25:7577–7586. - PubMed
-
- Yuan X., Zhou Y., Casanova E., Chai M., Kiss E., Grone H.J., Schutz G., Grummt I. Genetic inactivation of the transcription factor TIF-1A leads to nucleolar disruption, cell cycle arrest, and p53-mediated apoptosis. Mol. Cell. 2005;19:77–87. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
