

A monoclonal antibody specific for Candida albicans Als4 demonstrates overlapping localization of Als family proteins on the fungal cell surface and highlights differences between Als localization in vitro and in vivo
- PMID: 22106872
- PMCID: PMC3299873
- DOI: 10.1111/j.1574-695X.2011.00914.x
A monoclonal antibody specific for Candida albicans Als4 demonstrates overlapping localization of Als family proteins on the fungal cell surface and highlights differences between Als localization in vitro and in vivo
Abstract
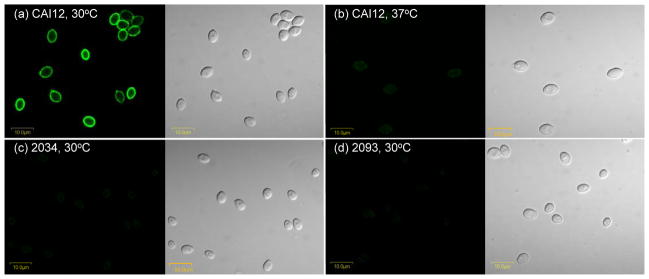
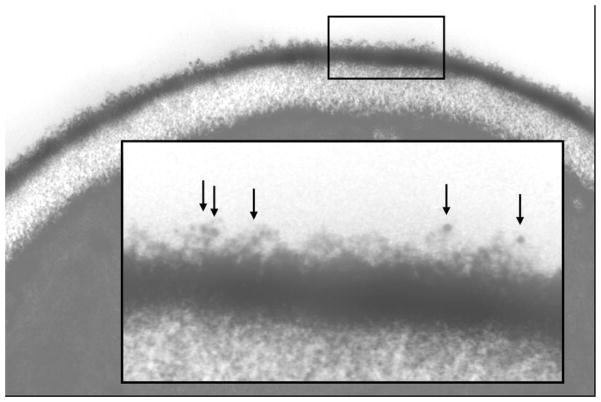
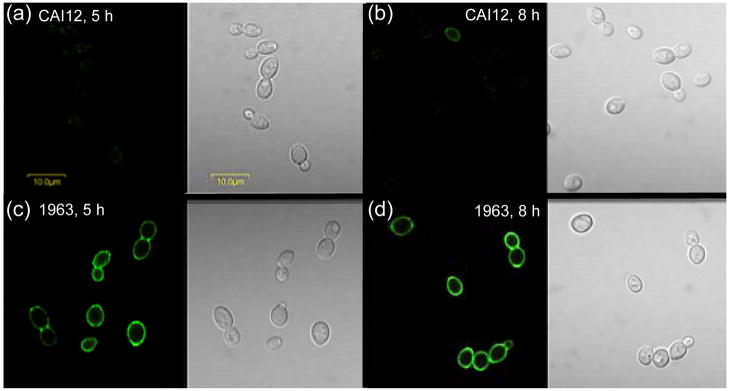
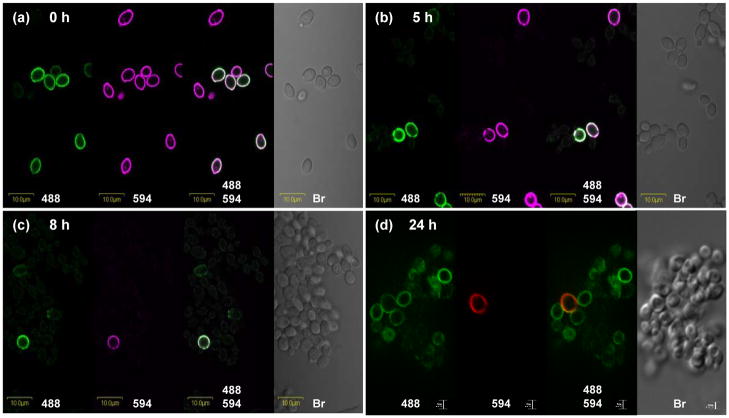
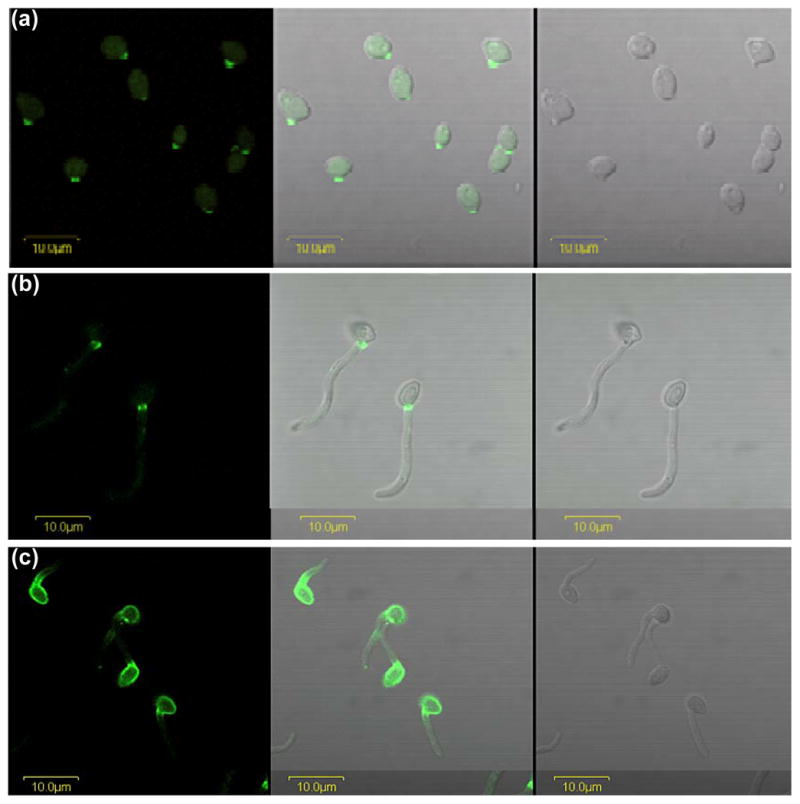
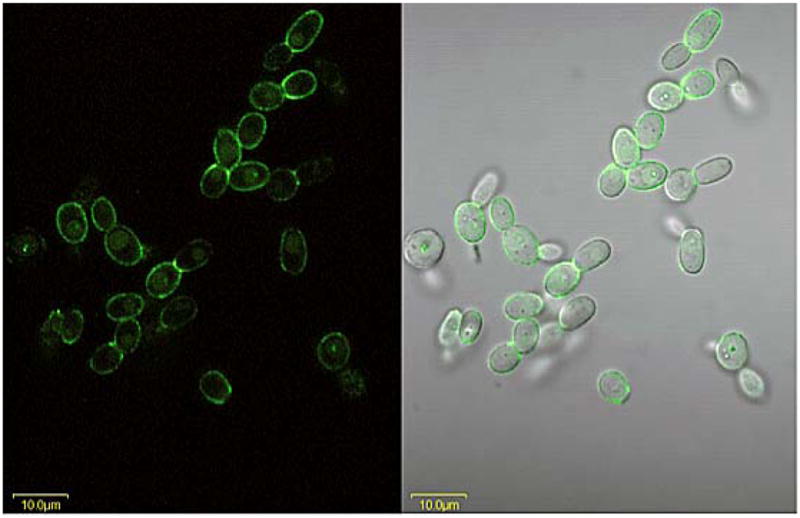
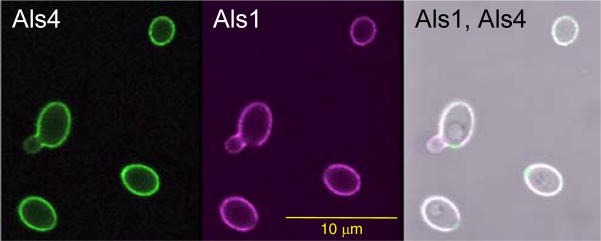
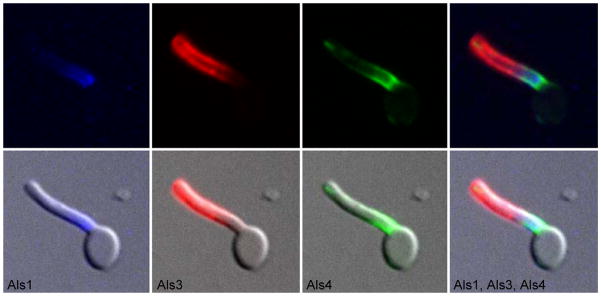
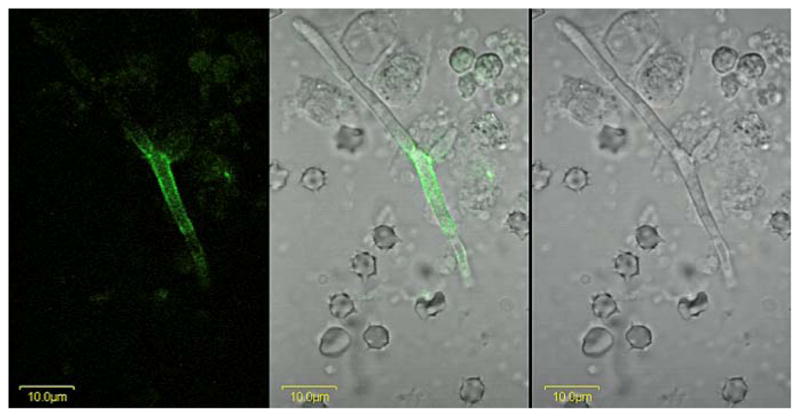
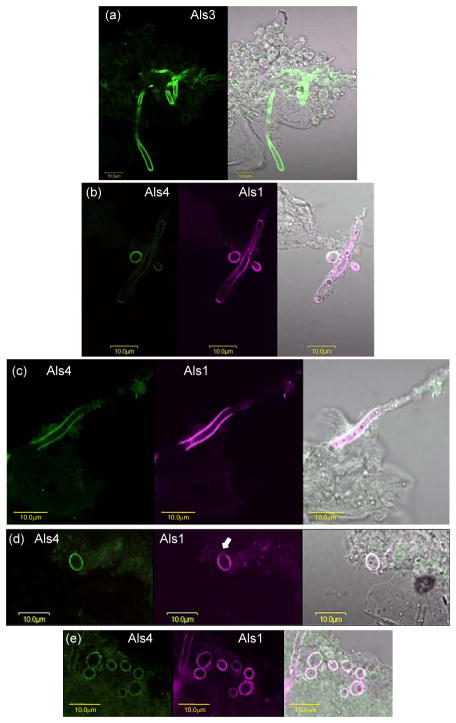
The Candida albicans agglutinin-like sequence (ALS) family encodes large cell surface glycoproteins that function in adhesion of the fungus to host and abiotic surfaces. Monoclonal antibodies (mAbs) specific for each Als protein were developed to study Als localization on the C. albicans surface. An anti-Als4 mAb demonstrated that Als4 covers the surface of yeast cells, with a greater abundance of Als4 on cells grown at 30 °C compared to 37 °C. On germ tubes, Als4 is localized in a restricted area proximal to the mother yeast. Immunolabeling with several anti-Als mAbs showed overlapping localization of Als1 and Als4 on yeast cells and Als1, Als3 and Als4 on germ tubes. Overlapping localization of Als proteins was also observed on yeast and hyphae recovered from mouse models of disseminated and oral candidiasis. Differences between Als localization in vivo and in vitro suggested changes in regulation of Als production in the host compared to the culture flask. Characterization with the anti-Als mAbs reveals the simultaneous presence and differences in relative abundance of Als proteins, creating an accurate image of Als representation and localization that can be used to guide conclusions regarding individual and collective Als protein function.
© 2011 Federation of European Microbiological Societies. Published by Blackwell Publishing Ltd. All rights reserved.
Figures
References
-
- Clancy CJ, Cheng S, Nguyen MH. Animal models of candidiasis. Methods Mol Biol. 2009;499:65–76. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous