

Differential effects of a polyalanine tract expansion in Arx on neural development and gene expression
- PMID: 22108177
- PMCID: PMC3277309
- DOI: 10.1093/hmg/ddr538
Differential effects of a polyalanine tract expansion in Arx on neural development and gene expression
Abstract
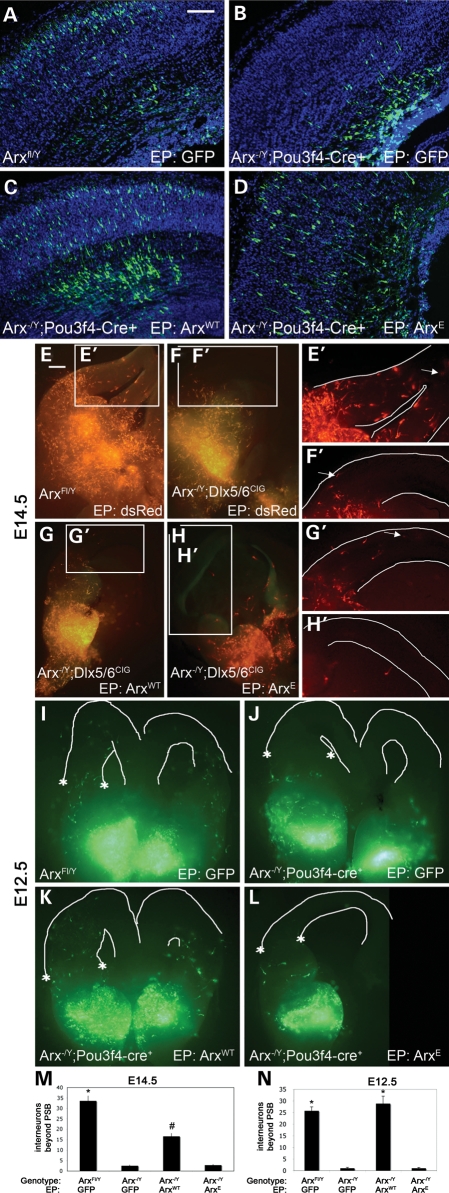
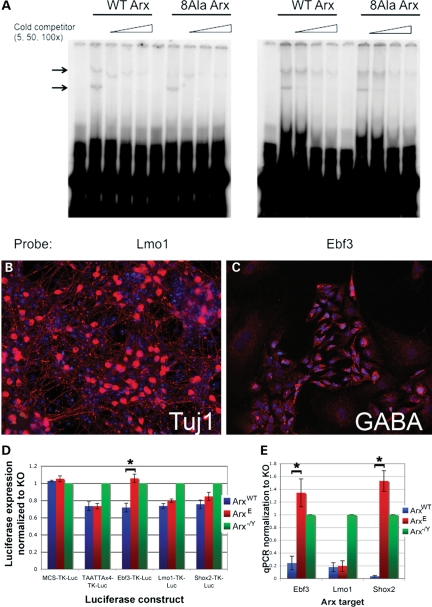
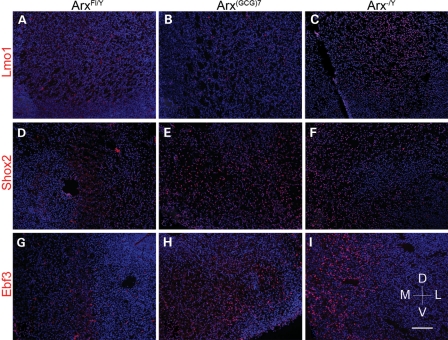
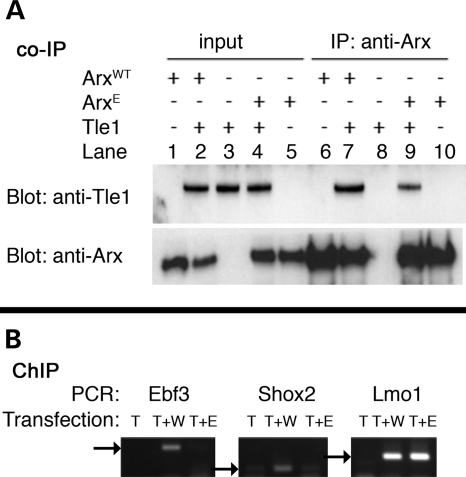
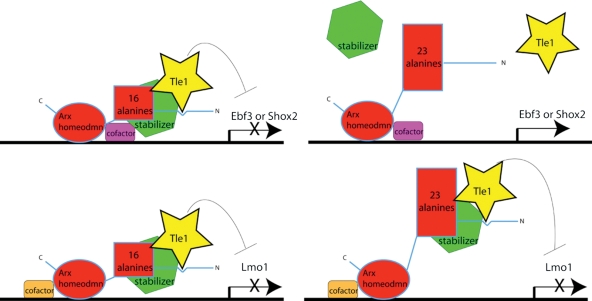
Polyalanine (poly-A) tracts exist in 494 annotated proteins; to date, expansions in these tracts have been associated with nine human diseases. The pathogenetic mechanism by which a poly-A tract results in these various human disorders remains uncertain. To understand the role of this mutation type, we investigated the change in functional properties of the transcription factor Arx when it has an expanded poly-A tract (Arx(E)), a mutation associated with infantile spasms and intellectual disabilities in humans. We found that although Arx(E) functions normally in the dorsal brain, its function in subpallial-derived populations of neurons is compromised. These contrasting functions are associated with the misregulation of Arx targets through the loss of the ability of Arx(E) to interact with the Arx cofactor Tle1. Our data demonstrate a novel mechanism for poly-A expansion diseases: the misregulation of a subset of target genes normally regulated by a transcription factor.
Figures
References
-
- Bienvenu T., Poirier K., Friocourt G., Bahi N., Beaumont D., Fauchereau F., Ben Jeema L., Zemni R., Vinet M.C., Francis F., et al. ARX, a novel Prd-class-homeobox gene highly expressed in the telencephalon, is mutated in X-linked mental retardation. Hum. Mol. Genet. 2002;11:981–991. - PubMed
-
- Guerrini R., Moro F., Kato M., Barkovich A.J., Shiihara T., McShane M.A., Hurst J., Loi M., Tohyama J., Norci V., et al. Expansion of the first PolyA tract of ARX causes infantile spasms and status dystonicus. Neurology. 2007;69:427–433. - PubMed
-
- Bauer P.O., Nukina N. The pathogenic mechanisms of polyglutamine diseases and current therapeutic strategies. J. Neurochem. 2009;110:1737–1765. - PubMed
-
- Messaed C., Rouleau G.A. Molecular mechanisms underlying polyalanine diseases. Neurobiol. Dis. 2009;34:397–405. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
