

Interplay between BRCA1 and RHAMM regulates epithelial apicobasal polarization and may influence risk of breast cancer
- PMID: 22110403
- PMCID: PMC3217025
- DOI: 10.1371/journal.pbio.1001199
Interplay between BRCA1 and RHAMM regulates epithelial apicobasal polarization and may influence risk of breast cancer
Abstract
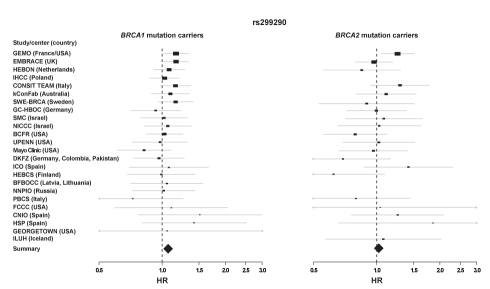
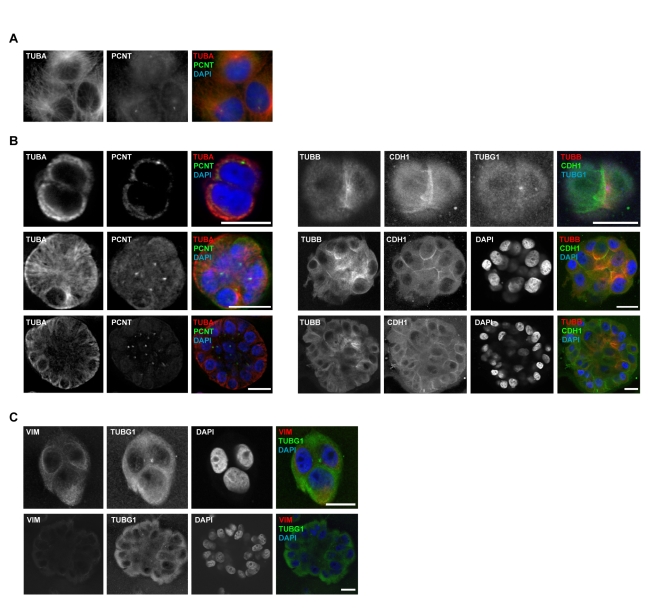
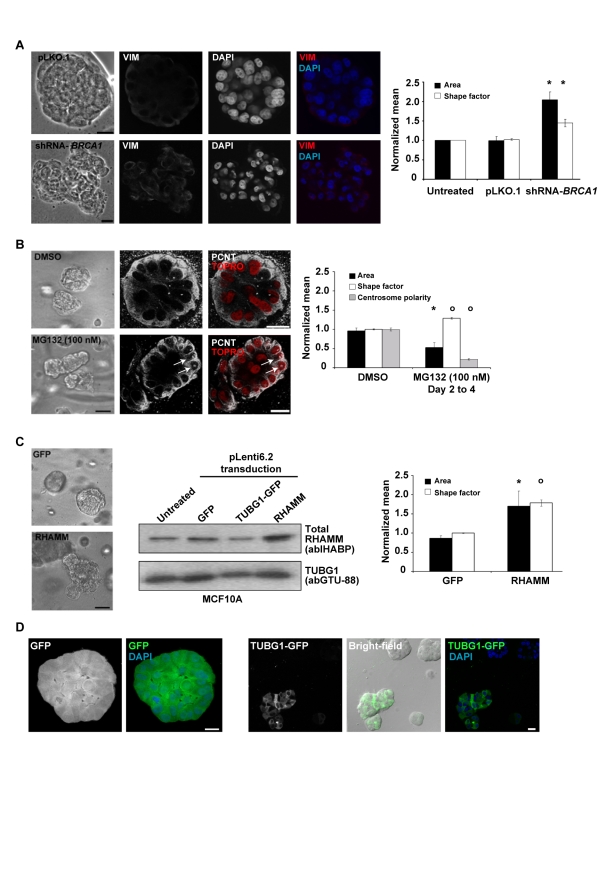
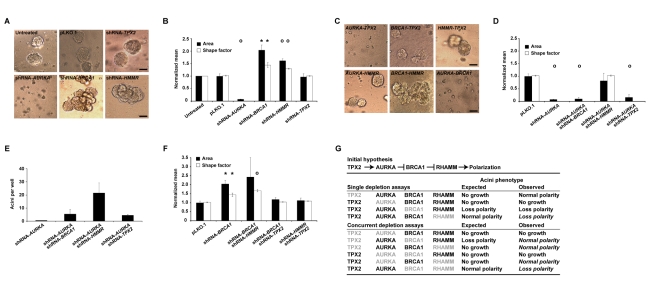
Differentiated mammary epithelium shows apicobasal polarity, and loss of tissue organization is an early hallmark of breast carcinogenesis. In BRCA1 mutation carriers, accumulation of stem and progenitor cells in normal breast tissue and increased risk of developing tumors of basal-like type suggest that BRCA1 regulates stem/progenitor cell proliferation and differentiation. However, the function of BRCA1 in this process and its link to carcinogenesis remain unknown. Here we depict a molecular mechanism involving BRCA1 and RHAMM that regulates apicobasal polarity and, when perturbed, may increase risk of breast cancer. Starting from complementary genetic analyses across families and populations, we identified common genetic variation at the low-penetrance susceptibility HMMR locus (encoding for RHAMM) that modifies breast cancer risk among BRCA1, but probably not BRCA2, mutation carriers: n = 7,584, weighted hazard ratio ((w)HR) = 1.09 (95% CI 1.02-1.16), p(trend) = 0.017; and n = 3,965, (w)HR = 1.04 (95% CI 0.94-1.16), p(trend) = 0.43; respectively. Subsequently, studies of MCF10A apicobasal polarization revealed a central role for BRCA1 and RHAMM, together with AURKA and TPX2, in essential reorganization of microtubules. Mechanistically, reorganization is facilitated by BRCA1 and impaired by AURKA, which is regulated by negative feedback involving RHAMM and TPX2. Taken together, our data provide fundamental insight into apicobasal polarization through BRCA1 function, which may explain the expanded cell subsets and characteristic tumor type accompanying BRCA1 mutation, while also linking this process to sporadic breast cancer through perturbation of HMMR/RHAMM.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
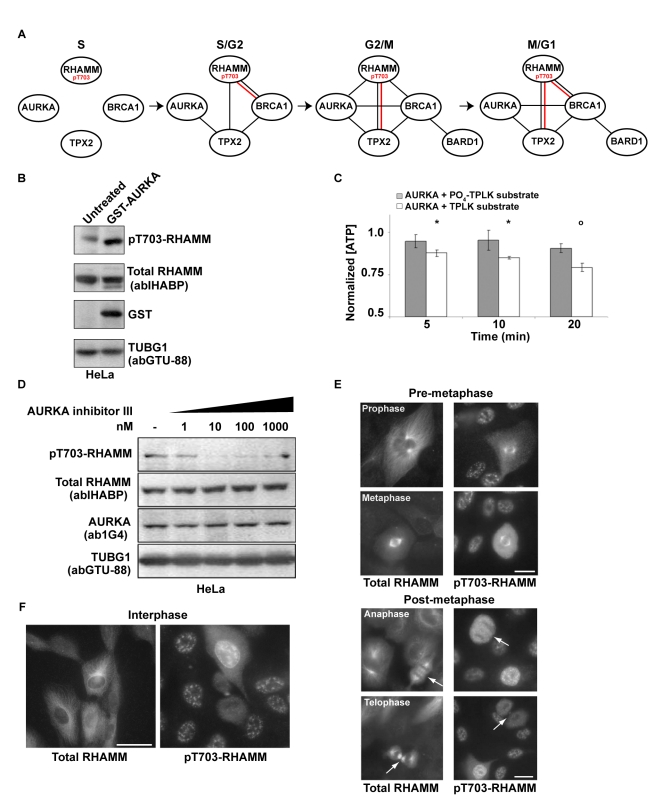
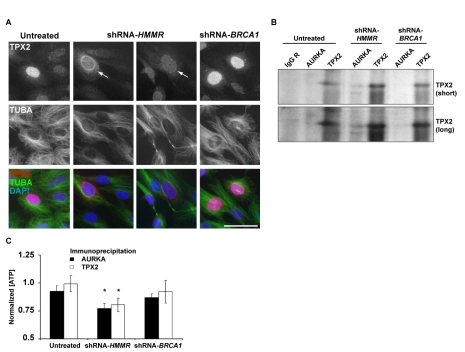
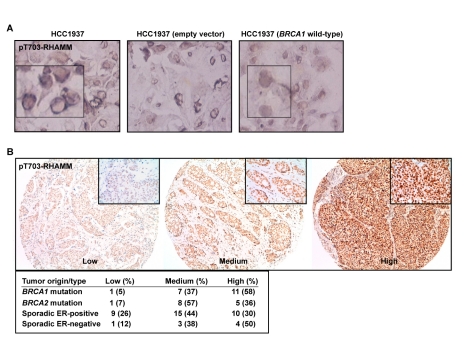
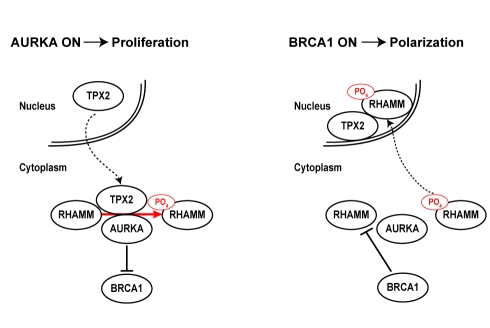
References
-
- Asselin-Labat M. L, Shackleton M, Stingl J, Vaillant F, Forrest N. C, et al. Steroid hormone receptor status of mouse mammary stem cells. J Natl Cancer Inst. 2006;98:1011–1014. - PubMed
-
- Shackleton M, Vaillant F, Simpson K. J, Stingl J, Smyth G. K, et al. Generation of a functional mammary gland from a single stem cell. Nature. 2006;439:84–88. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- U01 CA069417/CA/NCI NIH HHS/United States
- CA128978/CA/NCI NIH HHS/United States
- R25 CA092049/CA/NCI NIH HHS/United States
- 11174/CRUK_/Cancer Research UK/United Kingdom
- R01 CA128978/CA/NCI NIH HHS/United States
- CA69417/CA/NCI NIH HHS/United States
- U01 CA113916/CA/NCI NIH HHS/United States
- CA69638/CA/NCI NIH HHS/United States
- CA-06-503/CA/NCI NIH HHS/United States
- 10118/CRUK_/Cancer Research UK/United Kingdom
- CA69446/CA/NCI NIH HHS/United States
- U01 CA069638/CA/NCI NIH HHS/United States
- P50 CA83638/CA/NCI NIH HHS/United States
- CA69631/CA/NCI NIH HHS/United States
- U01 CA069467/CA/NCI NIH HHS/United States
- CA69467/CA/NCI NIH HHS/United States
- 11022/CRUK_/Cancer Research UK/United Kingdom
- CA122340/CA/NCI NIH HHS/United States
- U01 CA069398/CA/NCI NIH HHS/United States
- DH_/Department of Health/United Kingdom
- U01 CA069631/CA/NCI NIH HHS/United States
- U01 CA69398/CA/NCI NIH HHS/United States
- P50 CA083638/CA/NCI NIH HHS/United States
- R01 CA122340/CA/NCI NIH HHS/United States
- CA113916/CA/NCI NIH HHS/United States
- U01 CA069446/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
