

Fatal S. aureus hemorrhagic pneumonia: genetic analysis of a unique clinical isolate producing both PVL and TSST-1
- PMID: 22110621
- PMCID: PMC3207839
- DOI: 10.1371/journal.pone.0027246
Fatal S. aureus hemorrhagic pneumonia: genetic analysis of a unique clinical isolate producing both PVL and TSST-1
Abstract
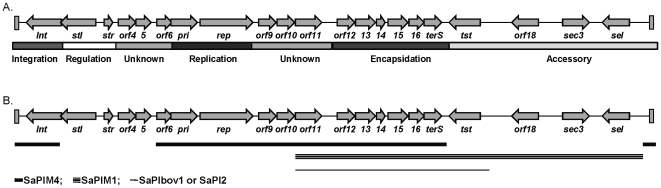
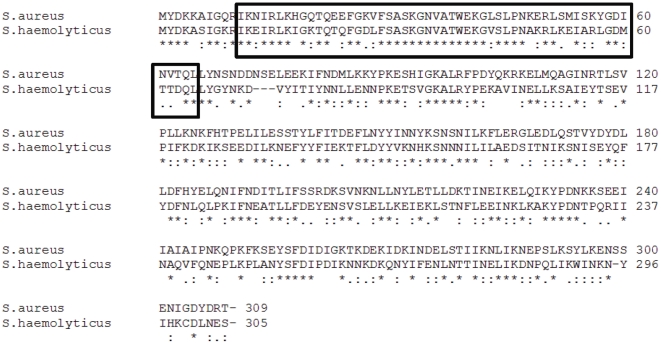
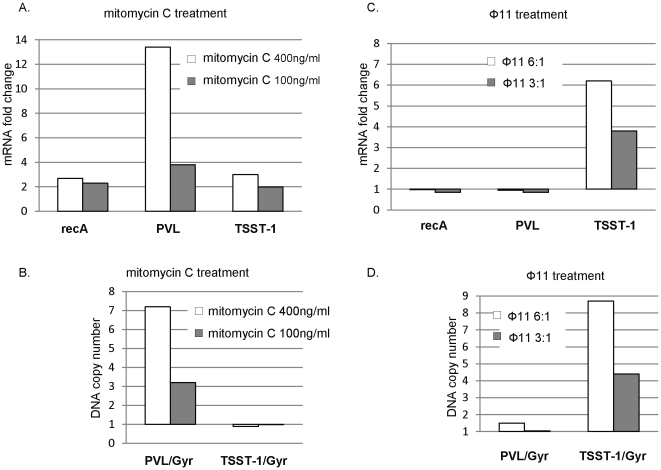
In 2008, an unusual strain of methicillin-sensitive Staphylococcus aureus (MSSA68111), producing both Panton-Valentine leukocidin (PVL) and toxic shock syndrome toxin-1 (TSST-1), was isolated from a fatal case of necrotizing pneumonia. Because PVL/TSST-1 co-production in S. aureus is rare, we characterized the molecular organization of these toxin genes in strain 68111. MSSA68111 carries the PVL genes within a novel temperate prophage we call ФPVLv68111 that is most similar, though not identical, to phage ФPVL--a phage type that is relatively rare worldwide. The TSST-1 gene (tst) in MSSA68111 is carried on a unique staphylococcal pathogenicity island (SaPI) we call SaPI68111. Features of SaPI68111 suggest it likely arose through multiple major recombination events with other known SaPIs. Both ФPVLv68111 and SaPI68111 are fully mobilizable and therefore transmissible to other strains. Taken together, these findings suggest that hypervirulent S. aureus have the potential to emerge worldwide.
Conflict of interest statement
Figures


References
-
- Archer GL. Staphylococcus aureus: a well-armed pathogen. Clin Infect Dis. 1998;26:1179–1181. - PubMed
-
- Lowy FD. Staphylococcus aureus infections. N Engl J Med. 1998;339:520–532. 10.1056/NEJM199808203390806 [doi] - PubMed
-
- Kaneko J, Kamio Y. Bacterial two-component and hetero-heptameric pore-forming cytolytic toxins: structures, pore-forming mechanism, and organization of the genes. Biosci Biotechnol Biochem. 2004;68:981–1003. - PubMed
-
- Gillet Y, Issartel B, Vanhems P, Fournet JC, Lina G, et al. Association between Staphylococcus aureus strains carrying gene for Panton-Valentine leukocidin and highly lethal necrotising pneumonia in young immunocompetent patients. Lancet. 2002;359:753–759. S0140-6736(02)07877-7 [pii];10.1016/S0140-6736(02)07877-7 [doi] - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
