

The identification of a small molecule compound that reduces HIV-1 Nef-mediated viral infectivity enhancement
- PMID: 22110726
- PMCID: PMC3217016
- DOI: 10.1371/journal.pone.0027696
The identification of a small molecule compound that reduces HIV-1 Nef-mediated viral infectivity enhancement
Abstract
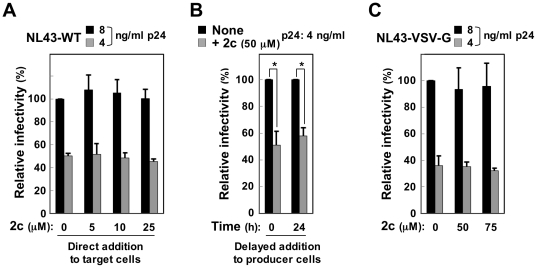
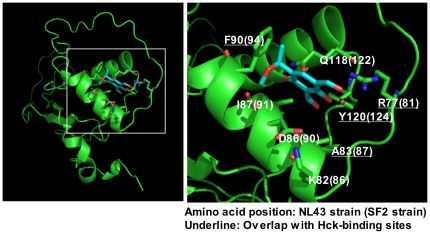
Nef is a multifunctional HIV-1 protein that accelerates progression to AIDS, and enhances the infectivity of progeny viruses through a mechanism that is not yet understood. Here, we show that the small molecule compound 2c reduces Nef-mediated viral infectivity enhancement. When added to viral producer cells, 2c did not affect the efficiency of viral production itself. However, the infectivity of the viruses produced in the presence of 2c was significantly lower than that of control viruses. Importantly, an inhibitory effect was observed with Nef(+) wild-type viruses, but not with viruses produced in the absence of Nef or in the presence of proline-rich PxxP motif-disrupted Nef, both of which displayed significantly reduced intrinsic infectivity. Meanwhile, the overexpression of the SH3 domain of the tyrosine kinase Hck, which binds to a PxxP motif in Nef, also reduced viral infectivity. Importantly, 2c inhibited Hck SH3-Nef binding, which was more marked when Nef was pre-incubated with 2c prior to its incubation with Hck, indicating that both Hck SH3 and 2c directly bind to Nef and that their binding sites overlap. These results imply that both 2c and the Hck SH3 domain inhibit the interaction of Nef with an unidentified host protein and thereby reduce Nef-mediated infectivity enhancement. The first inhibitory compound 2c is therefore a valuable chemical probe for revealing the underlying molecular mechanism by which Nef enhances the infectivity of HIV-1.
Conflict of interest statement
Figures








Similar articles
-
Interaction with the Src homology (SH3-SH2) region of the Src-family kinase Hck structures the HIV-1 Nef dimer for kinase activation and effector recruitment.J Biol Chem. 2014 Oct 10;289(41):28539-53. doi: 10.1074/jbc.M114.600031. Epub 2014 Aug 13. J Biol Chem. 2014. PMID: 25122770 Free PMC article.
-
HIV-1 infection of T cells and macrophages are differentially modulated by virion-associated Hck: a Nef-dependent phenomenon.Viruses. 2013 Sep 18;5(9):2235-52. doi: 10.3390/v5092235. Viruses. 2013. PMID: 24051604 Free PMC article.
-
The cellular kinase binding motifs (PxxP and RR) in human immunodeficiency virus type 1 Nef protein are dispensable for producer-cell-dependent enhancement of viral entry.Virology. 1999 May 10;257(2):285-9. doi: 10.1006/viro.1999.9682. Virology. 1999. PMID: 10329538
-
Structure, function, and inhibitor targeting of HIV-1 Nef-effector kinase complexes.J Biol Chem. 2020 Oct 30;295(44):15158-15171. doi: 10.1074/jbc.REV120.012317. Epub 2020 Aug 29. J Biol Chem. 2020. PMID: 32862141 Free PMC article. Review.
-
Hck inhibitors as potential therapeutic agents in cancer and HIV infection.Curr Med Chem. 2015;22(13):1540-64. doi: 10.2174/0929867322666150209152057. Curr Med Chem. 2015. PMID: 25666803 Review.
Cited by
-
Open-channel blocking action of volatile anaesthetics desflurane and sevoflurane on human voltage-gated Kv 1.5 channel.Br J Pharmacol. 2020 Aug;177(16):3811-3827. doi: 10.1111/bph.15105. Epub 2020 Jun 30. Br J Pharmacol. 2020. PMID: 32436224 Free PMC article.
-
Development and validation of a high-content bimolecular fluorescence complementation assay for small-molecule inhibitors of HIV-1 Nef dimerization.J Biomol Screen. 2014 Apr;19(4):556-65. doi: 10.1177/1087057113513640. Epub 2013 Nov 26. J Biomol Screen. 2014. PMID: 24282155 Free PMC article.
-
Overlapping effector interfaces define the multiple functions of the HIV-1 Nef polyproline helix.Retrovirology. 2012 May 31;9:47. doi: 10.1186/1742-4690-9-47. Retrovirology. 2012. PMID: 22651890 Free PMC article.
-
Small molecule inhibitors of the HIV-1 virulence factor, Nef.Drug Discov Today Technol. 2013 Dec;10(4):e523-9. doi: 10.1016/j.ddtec.2013.07.002. Drug Discov Today Technol. 2013. PMID: 24451644 Free PMC article. Review.
-
Antiretroviral Drug Discovery Targeting the HIV-1 Nef Virulence Factor.Viruses. 2022 Sep 13;14(9):2025. doi: 10.3390/v14092025. Viruses. 2022. PMID: 36146831 Free PMC article. Review.
References
-
- Fackler OT, Baur AS. Live and let die: Nef functions beyond HIV replication. Immunity. 2002;16:493–497. - PubMed
-
- Peterlin BM, Trono D. Hide, shield and strike back: how HIV-infected cells avoid immune eradication. Nat Rev Immunol. 2003;3:97–107. - PubMed
-
- Malim MH, Emerman M. HIV-1 accessory proteins–ensuring viral survival in a hostile environment. Cell Host Microbe. 2008;3:388–398. - PubMed
-
- Deacon NJ, Tsykin A, Solomon A, Smith K, Ludford-Menting M, et al. Genomic structure of an attenuated quasi species of HIV-1 from a blood transfusion donor and recipients. Science. 1995;270:988–991. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
