

Epigenetics and genetics. MicroRNAs en route to the clinic: progress in validating and targeting microRNAs for cancer therapy
- PMID: 22113163
- PMCID: PMC4314215
- DOI: 10.1038/nrc3166
Epigenetics and genetics. MicroRNAs en route to the clinic: progress in validating and targeting microRNAs for cancer therapy
Abstract
In normal cells multiple microRNAs (miRNAs) converge to maintain a proper balance of various processes, including proliferation, differentiation and cell death. miRNA dysregulation can have profound cellular consequences, especially because individual miRNAs can bind to and regulate multiple mRNAs. In cancer, the loss of tumour-suppressive miRNAs enhances the expression of target oncogenes, whereas increased expression of oncogenic miRNAs (known as oncomirs) can repress target tumour suppressor genes. This realization has resulted in a quest to understand the pathways that are regulated by these miRNAs using in vivo model systems, and to comprehend the feasibility of targeting oncogenic miRNAs and restoring tumour-suppressive miRNAs for cancer therapy. Here we discuss progress in using mouse models to understand the roles of miRNAs in cancer and the potential for manipulating miRNAs for cancer therapy as these molecules make their way towards clinical trials.
Figures
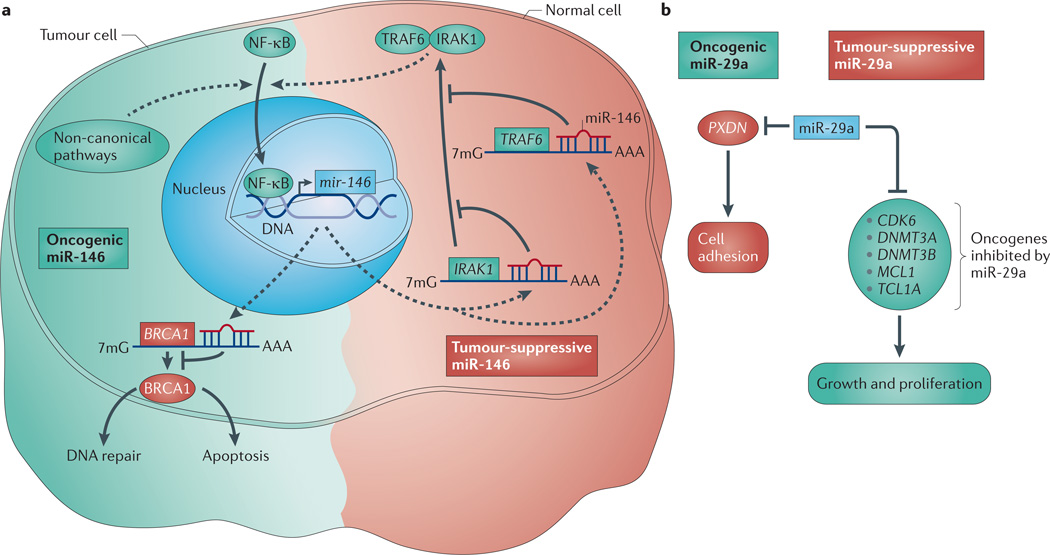
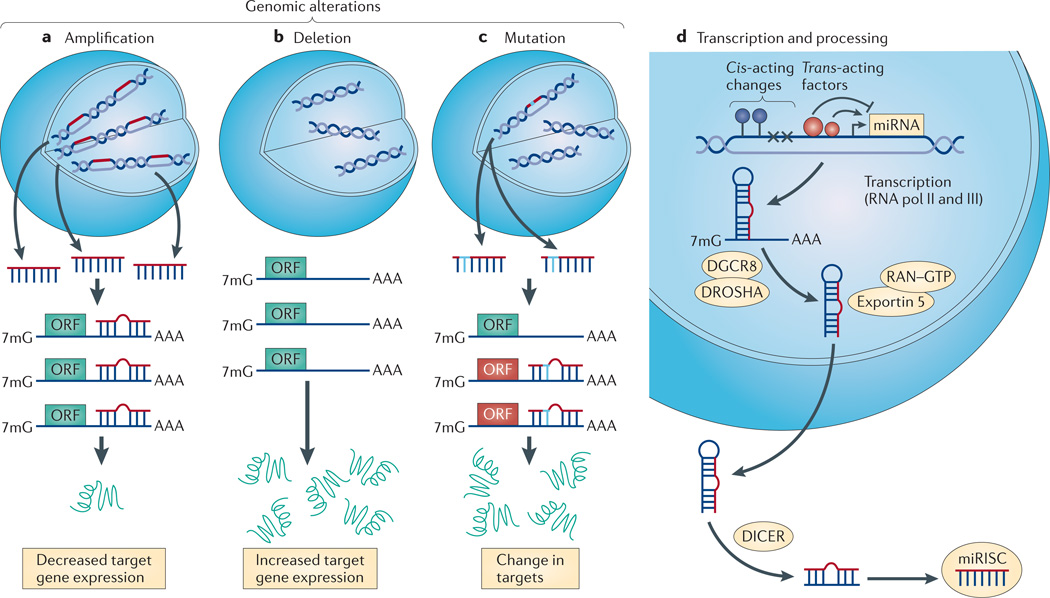
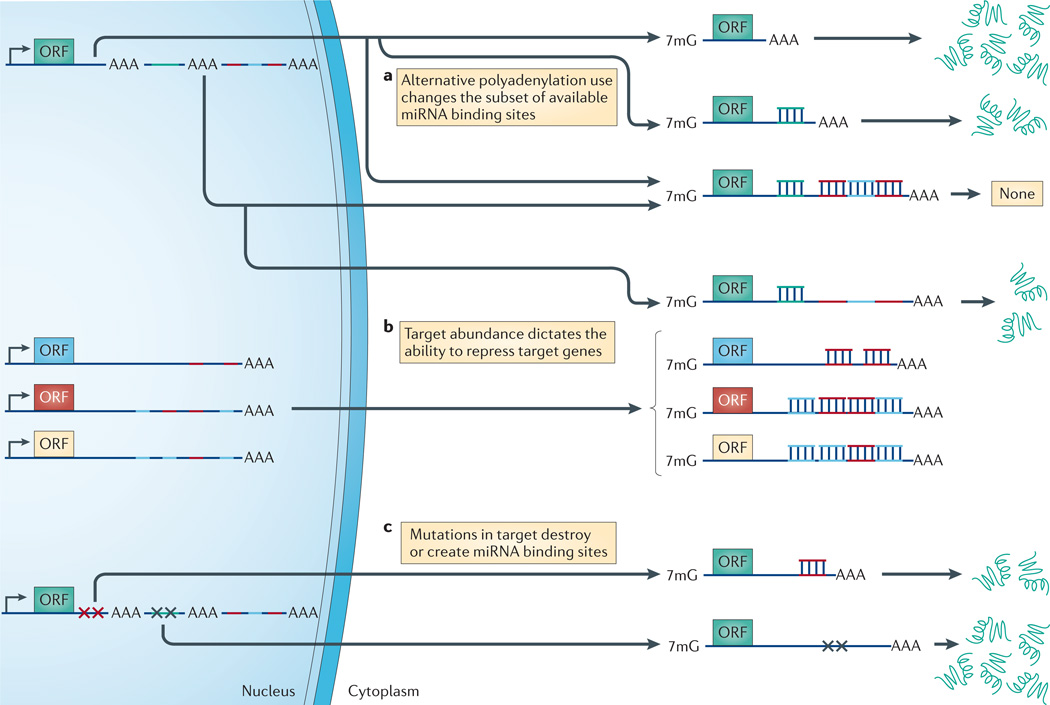
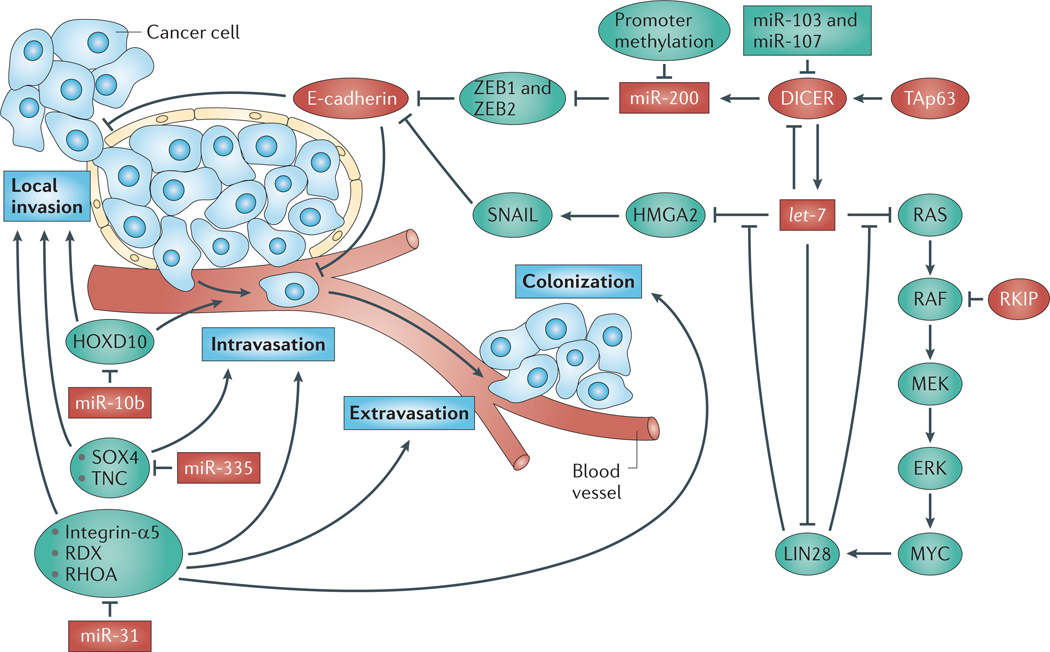
References
-
- Chalfie M, Horvitz HR, Sulston JE. Mutations that lead to reiterations in the cell lineages of C. elegans. Cell. 1981;24:59–69. - PubMed
-
- Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. 1993;75:843–854. - PubMed
-
- Reinhart BJ, et al. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature. 2000;403:901–906. - PubMed
-
- Slack FJ, et al. The lin-41 RBCC gene acts in the C. elegans heterochronic pathway between the let-7 regulatory RNA and the LIN-29 transcription factor. Mol. Cell. 2000;5:659–669. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
