

Evaluating genetic markers and neurobiochemical analytes for fluoxetine response using a panel of mouse inbred strains
- PMID: 22113448
- PMCID: PMC3337404
- DOI: 10.1007/s00213-011-2574-z
Evaluating genetic markers and neurobiochemical analytes for fluoxetine response using a panel of mouse inbred strains
Abstract
Rationale: Identification of biomarkers that establish diagnosis or treatment response is critical to the advancement of research and management of patients with depression.
Objective: Our goal was to identify biomarkers that can potentially assess fluoxetine response and risk to poor treatment outcome.
Methods: We measured behavior, gene expression, and the levels of 36 neurobiochemical analytes across a panel of genetically diverse mouse inbred lines after chronic treatment with water or fluoxetine.
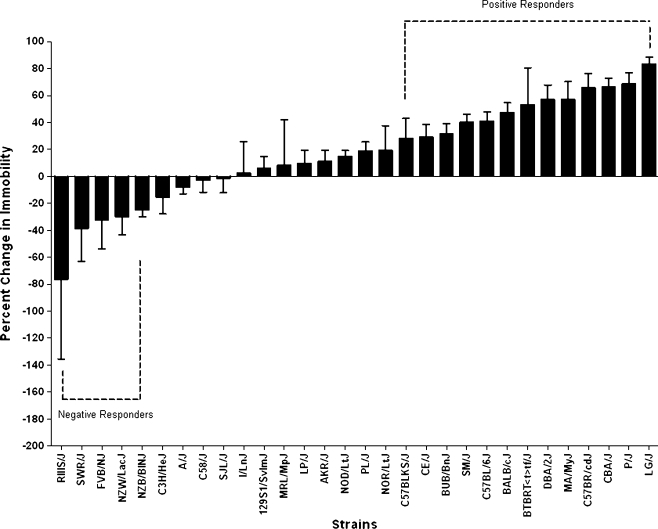
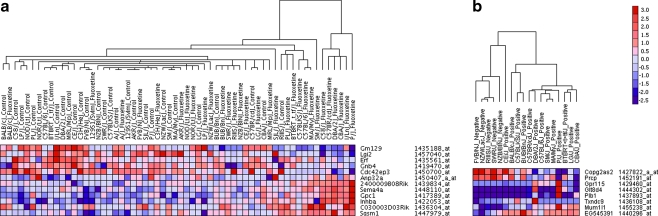
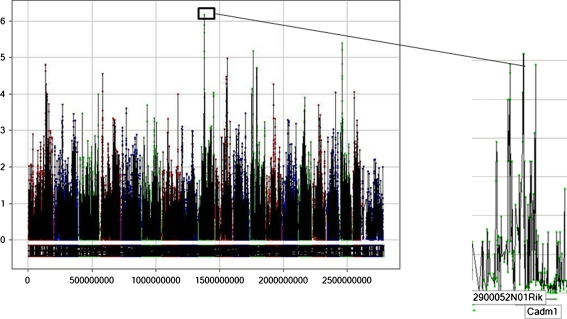
Results: Glyoxylase 1 (GLO1) and guanine nucleotide-binding protein 1 (GNB1) mostly account for baseline anxiety-like and depressive-like behavior, indicating a common biological link between depression and anxiety. Fluoxetine-induced biochemical alterations discriminated positive responders, while baseline neurobiochemical differences differentiated negative responders (p < 0.006). Results show that glial fibrillary acidic protein, S100 beta protein, GLO1, and histone deacetylase 5 contributed most to fluoxetine response. These proteins are linked within a cellular growth/proliferation pathway, suggesting the involvement of cellular genesis in fluoxetine response. Furthermore, a candidate genetic locus that associates with baseline depressive-like behavior contains a gene that encodes for cellular proliferation/adhesion molecule (Cadm1), supporting a genetic basis for the role of neuro/gliogenesis in depression.
Conclusion: We provided a comprehensive analysis of behavioral, neurobiochemical, and transcriptome data across 30 mouse inbred strains that has not been accomplished before. We identified biomarkers that influence fluoxetine response, which, altogether, implicate the importance of cellular genesis in fluoxetine treatment. More broadly, this approach can be used to assess a wide range of drug response phenotypes that are challenging to address in human samples.
Figures

References
-
- Amsterdam JD, Fawcett J, Quitkin FM, Reimherr FW, Rosenbaum JF, Michelson D, Hornig-Rohan M, Beasley CM. Fluoxetine and norfluoxetine plasma concentrations in major depression: a multicenter study. Am J Psychiatry. 1997;154:963–969. - PubMed
-
- Andreescu C, Lenze EJ, Dew MA, Begley AE, Mulsant BH, Dombrovski AY, Pollock BG, Stack J, Miller MD, Reynolds CF. Effect of comorbid anxiety on treatment response and relapse risk in late-life depression: Controlled study. Br J Psychiatry. 2007;190:344–349. doi: 10.1192/bjp.bp.106.027169. - DOI - PubMed
-
- Arai M, Yuzawa H, Nohara I, Ohnishi T, Obata N, Iwayama Y, Haga S, Toyota T, Ujike H, Ichikawa T, Nishida A, Tanaka Y, Furukawa A, Aikawa Y, Kuroda O, Niizato K, Izawa R, Nakamura K, Mori N, Matsuzawa D, Hashimoto K, Iyo M, Sora I, Matsushita M, Okazaki Y, Yoshikawa T, Miyata T, Itokawa M. Enhanced carbonyl stress in a subpopulation of schizophrenia. Arch Gen Psychiatry. 2010;67:589–597. doi: 10.1001/archgenpsychiatry.2010.62. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
