

Dual γ rhythm generators control interlaminar synchrony in auditory cortex
- PMID: 22114273
- PMCID: PMC3654396
- DOI: 10.1523/JNEUROSCI.2209-11.2011
Dual γ rhythm generators control interlaminar synchrony in auditory cortex
Erratum in
- J Neurosci. 2012 Feb 22;32(8):2911
Abstract
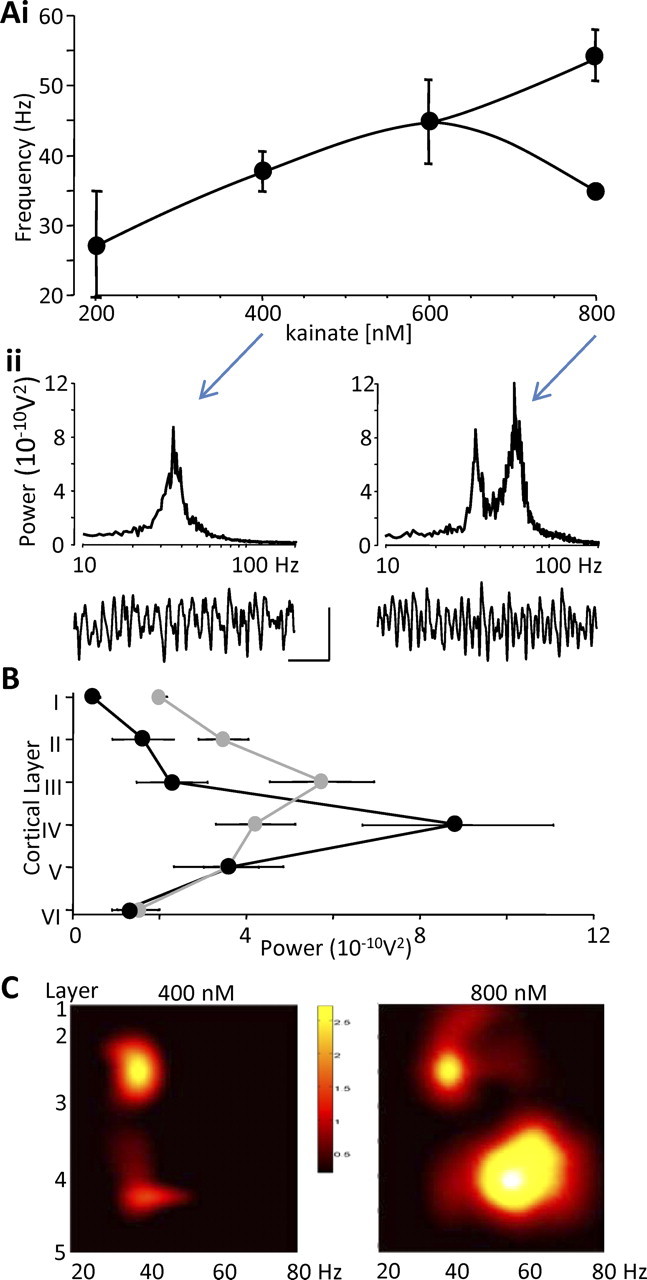
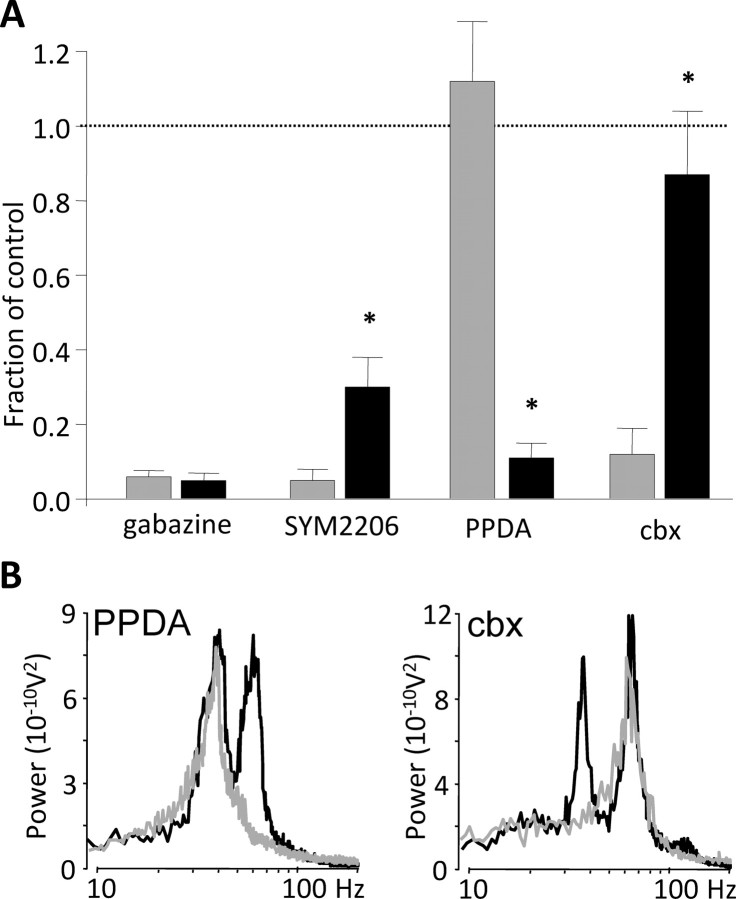
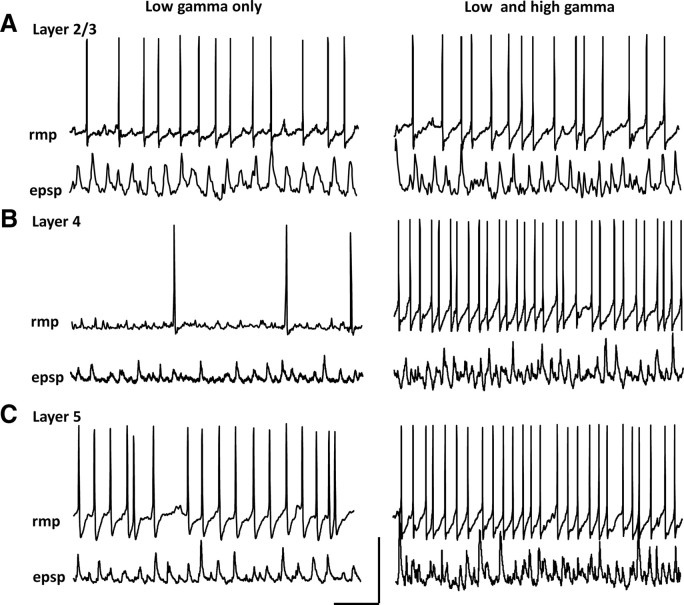
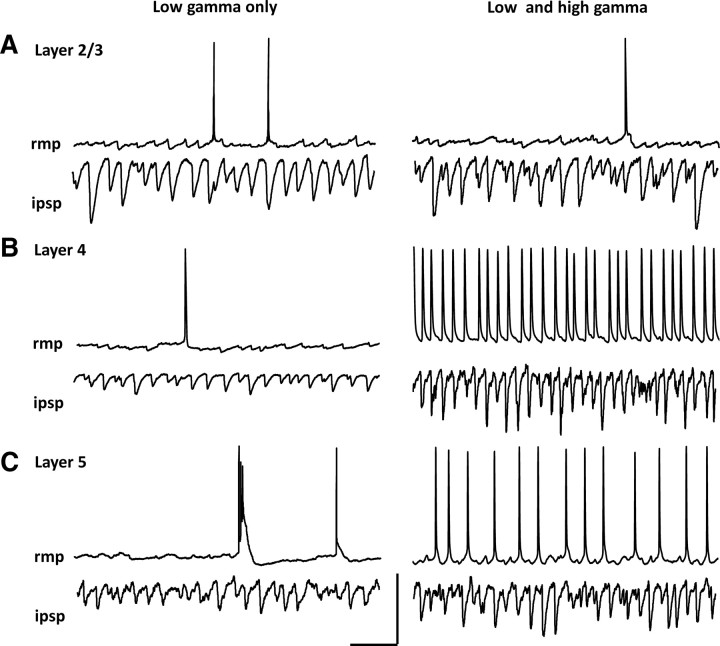
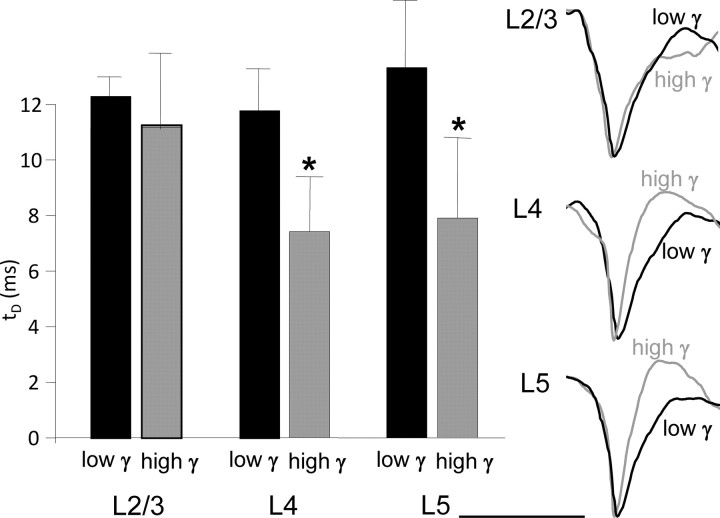
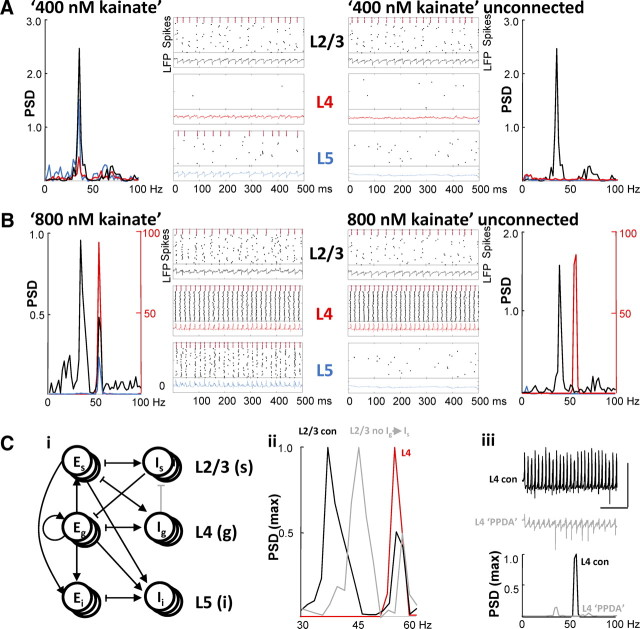
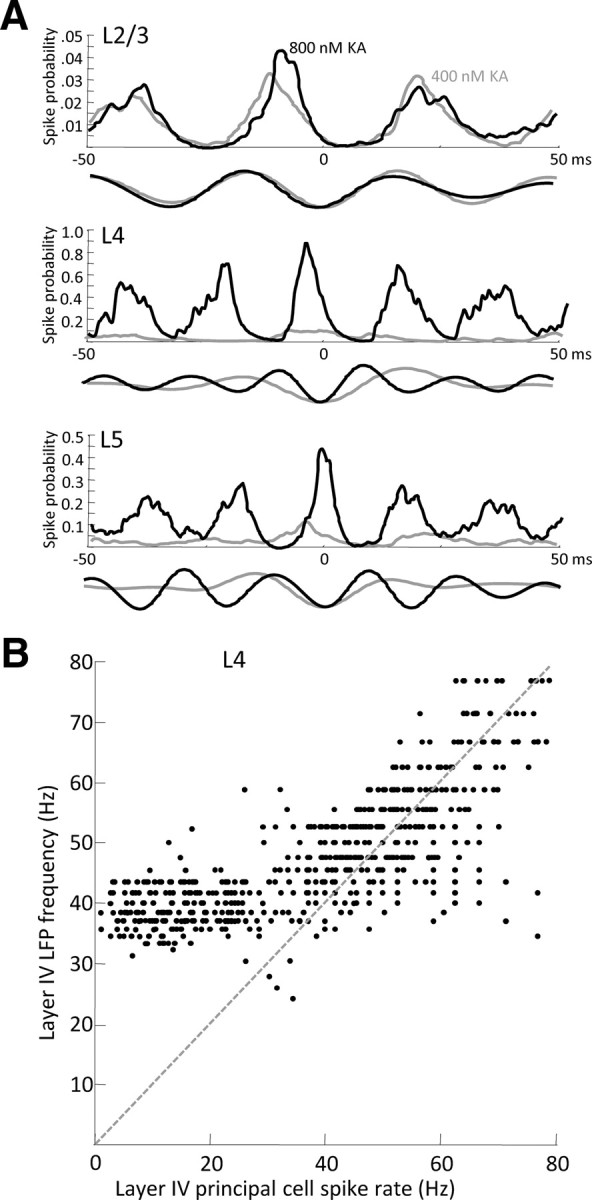
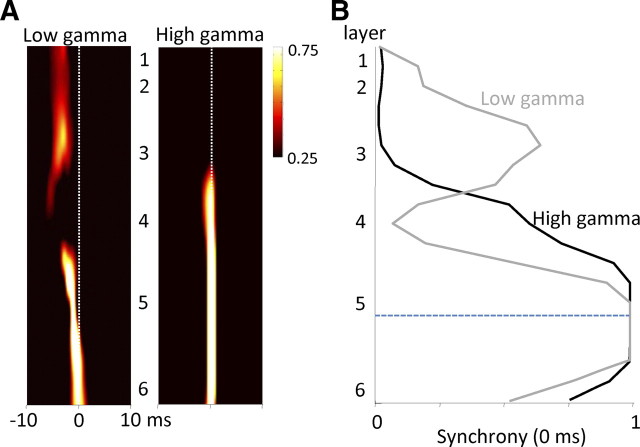
Rhythmic activity in populations of cortical neurons accompanies, and may underlie, many aspects of primary sensory processing and short-term memory. Activity in the gamma band (30 Hz up to >100 Hz) is associated with such cognitive tasks and is thought to provide a substrate for temporal coupling of spatially separate regions of the brain. However, such coupling requires close matching of frequencies in co-active areas, and because the nominal gamma band is so spectrally broad, it may not constitute a single underlying process. Here we show that, for inhibition-based gamma rhythms in vitro in rat neocortical slices, mechanistically distinct local circuit generators exist in different laminae of rat primary auditory cortex. A persistent, 30-45 Hz, gap-junction-dependent gamma rhythm dominates rhythmic activity in supragranular layers 2/3, whereas a tonic depolarization-dependent, 50-80 Hz, pyramidal/interneuron gamma rhythm is expressed in granular layer 4 with strong glutamatergic excitation. As a consequence, altering the degree of excitation of the auditory cortex causes bifurcation in the gamma frequency spectrum and can effectively switch temporal control of layer 5 from supragranular to granular layers. Computational modeling predicts the pattern of interlaminar connections may help to stabilize this bifurcation. The data suggest that different strategies are used by primary auditory cortex to represent weak and strong inputs, with principal cell firing rate becoming increasingly important as excitation strength increases.
Figures
References
-
- Compte A, Sanchez-Vives MV, McCormick DA, Wang XJ. Cellular and network mechanisms of slow oscillatory activity (<1 Hz) and wave propagations in a cortical network model. J Neurophysiol. 2003;89:2707–2725. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous