

Sodium-dependent vitamin C transporter 2 deficiency causes hypomyelination and extracellular matrix defects in the peripheral nervous system
- PMID: 22114285
- PMCID: PMC6623846
- DOI: 10.1523/JNEUROSCI.3457-11.2011
Sodium-dependent vitamin C transporter 2 deficiency causes hypomyelination and extracellular matrix defects in the peripheral nervous system
Abstract
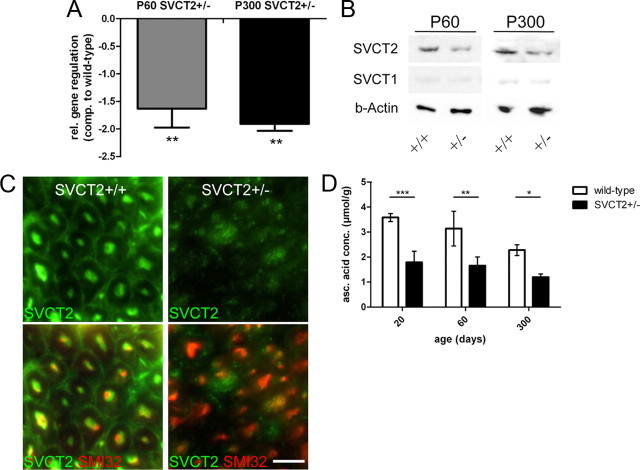
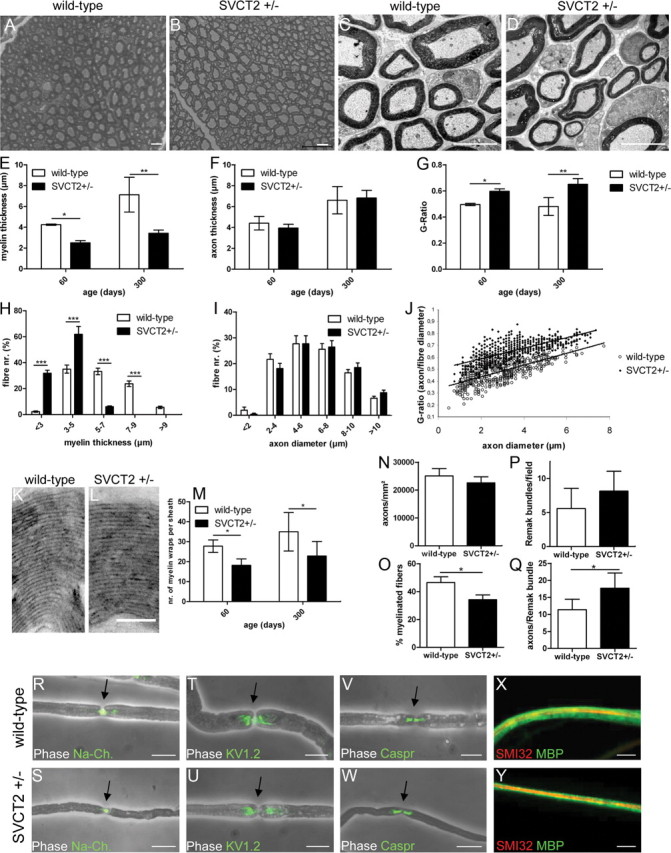
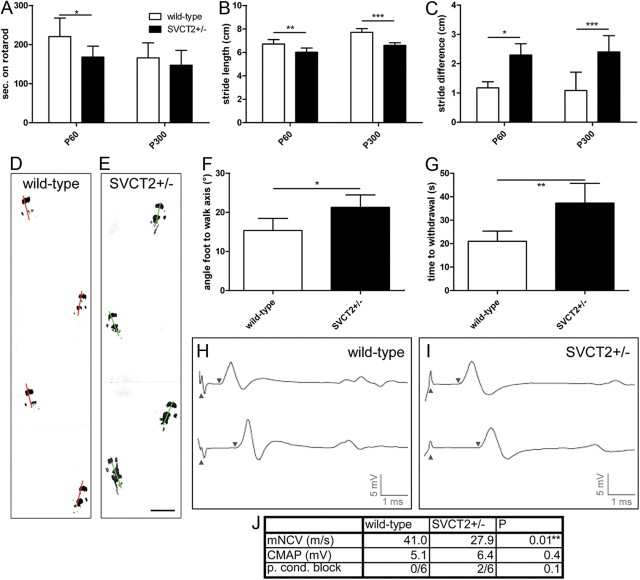
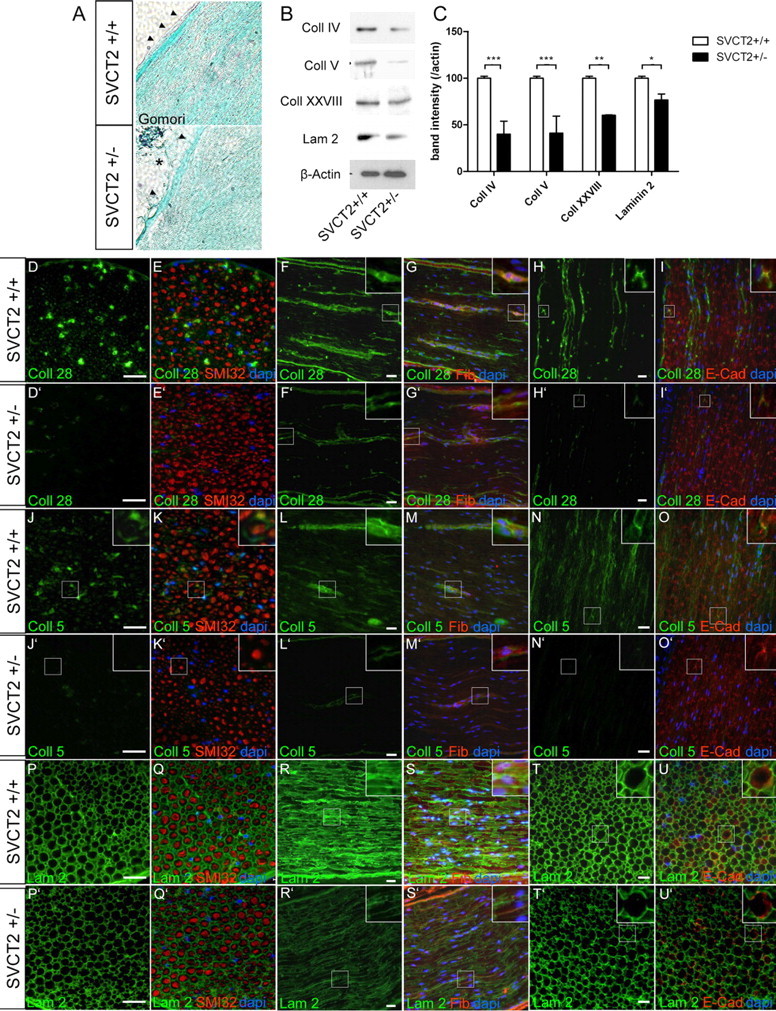
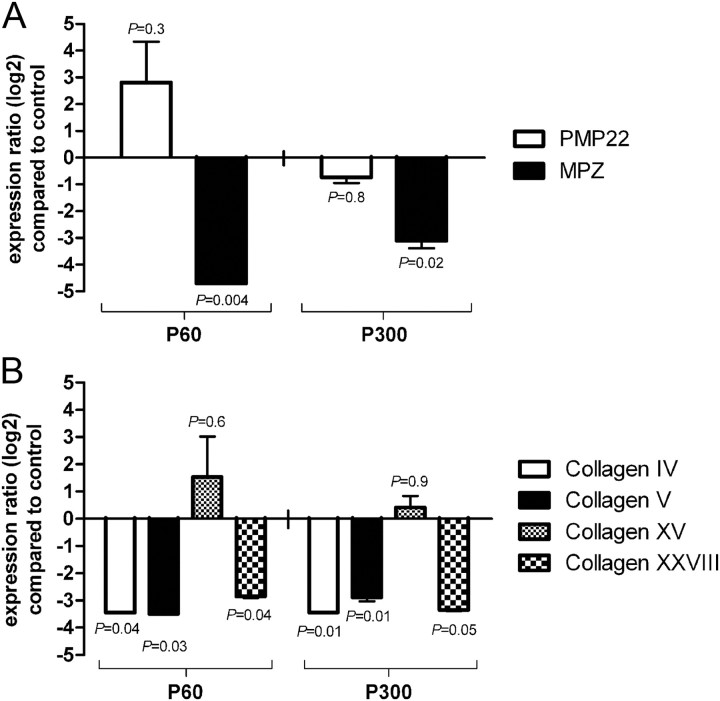
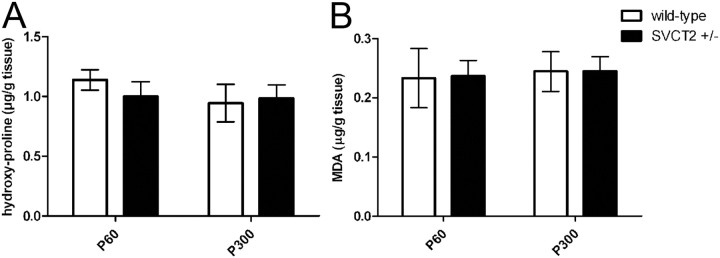
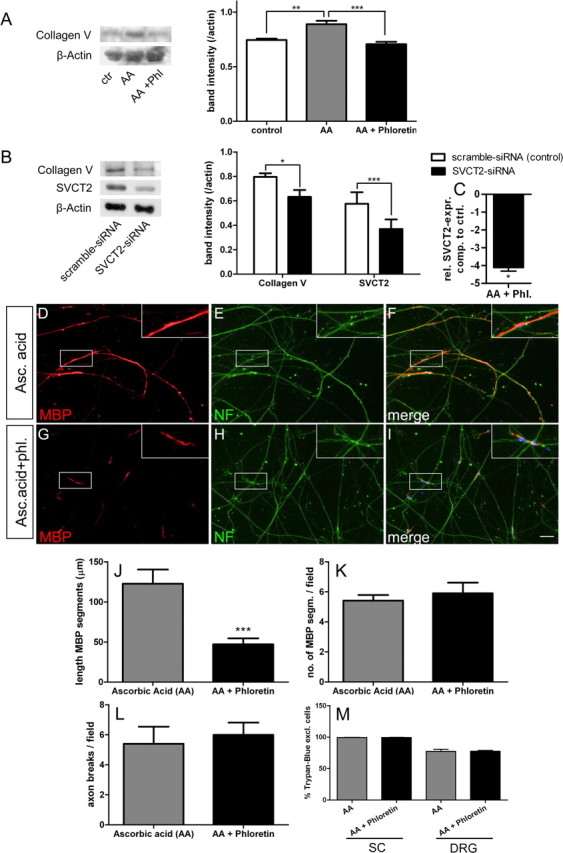
Ascorbic acid (vitamin C) is necessary for myelination of Schwann cell/neuron cocultures and has shown beneficial effects in the treatment of a Charcot-Marie-Tooth neuropathy 1A (CMT1A) mouse model. Although clinical studies revealed that ascorbic acid treatment had no impact on CMT1A, it is assumed to have an important function in peripheral nerve myelination and possibly in remyelination. However, the transport pathway of ascorbic acid into peripheral nerves and the mechanism of ascorbic acid function in peripheral nerves in vivo remained unclear. In this study, we used sodium-dependent vitamin C transporter 2-heterozygous (SVCT2(+/-)) mice to elucidate the functions of SVCT2 and ascorbic acid in the murine peripheral nervous system. SVCT2 and ascorbic acid levels were reduced in SVCT2(+/-) peripheral nerves. Morphometry of sciatic nerve fibers revealed a decrease in myelin thickness and an increase in G-ratios in SVCT2(+/-) mice. Nerve conduction velocities and sensorimotor performance in functional tests were reduced in SVCT2(+/-) mice. To investigate the mechanism of ascorbic acid function, we studied the expression of collagens in the extracellular matrix of peripheral nerves. Here, we show that expression of various collagen types was reduced in sciatic nerves of SVCT2(+/-) mice. We found that collagen gene transcription was reduced in SVCT2(+/-) mice but hydroxyproline levels were not, indicating that collagen formation was regulated on the transcriptional and not the posttranslational level. These results help to clarify the transport pathway and mechanism of action of ascorbic acid in the peripheral nervous system and may lead to novel therapeutic approaches to peripheral neuropathies by manipulation of SVCT2 function.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- Angelow S, Haselbach M, Galla HJ. Functional characterisation of the active ascorbic acid transport into cerebrospinal fluid using primary cultured choroid plexus cells. Brain Res. 2003;988:105–113. - PubMed
-
- Badcock NR, Zoanetti GD, Martin ES. Nonchromatographic assay for malondialdehyde-thiobarbituric acid adduct with HPLC equivalence. Clin Chem. 1997;43:1655–1657. - PubMed
-
- Bella J, Eaton M, Brodsky B, Berman HM. Crystal and molecular structure of a collagen-like peptide at 1.9 Å resolution. Science. 1994;266:75–81. - PubMed
-
- Booth BA, Uitto J. Collagen biosynthesis by human skin fibroblasts. III. The effects of ascorbic acid on procollagen production and prolyl hydroxylase activity. Biochim Biophys Acta. 1981;675:117–122. - PubMed
-
- Burns J, Ouvrier RA, Yiu EM, Joseph PD, Kornberg AJ, Fahey MC, Ryan MM. Ascorbic acid for Charcot-Marie-Tooth disease type 1A in children: a randomised, double-blind, placebo-controlled, safety and efficacy trial. Lancet Neurol. 2009;8:537–544. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases