

A TNF-regulated recombinatorial macrophage immune receptor implicated in granuloma formation in tuberculosis
- PMID: 22114556
- PMCID: PMC3219713
- DOI: 10.1371/journal.ppat.1002375
A TNF-regulated recombinatorial macrophage immune receptor implicated in granuloma formation in tuberculosis
Abstract
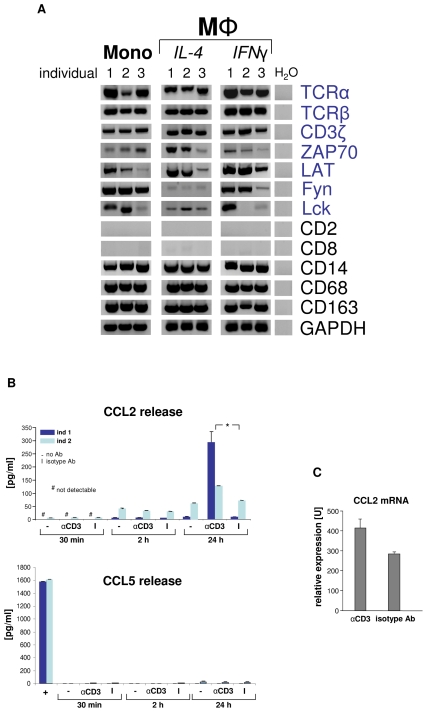
Macrophages play a central role in host defense against mycobacterial infection and anti- TNF therapy is associated with granuloma disorganization and reactivation of tuberculosis in humans. Here, we provide evidence for the presence of a T cell receptor (TCR) αβ based recombinatorial immune receptor in subpopulations of human and mouse monocytes and macrophages. In vitro, we find that the macrophage-TCRαβ induces the release of CCL2 and modulates phagocytosis. TNF blockade suppresses macrophage-TCRαβ expression. Infection of macrophages from healthy individuals with mycobacteria triggers formation of clusters that express restricted TCR Vβ repertoires. In vivo, TCRαβ bearing macrophages abundantly accumulate at the inner host-pathogen contact zone of caseous granulomas from patients with lung tuberculosis. In chimeric mouse models, deletion of the variable macrophage-TCRαβ or TNF is associated with structurally compromised granulomas of pulmonary tuberculosis even in the presence of intact T cells. These results uncover a TNF-regulated recombinatorial immune receptor in monocytes/macrophages and demonstrate its implication in granuloma formation in tuberculosis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures








References
-
- Janeway CA, Jr, Medzhitov R. Innate immune recognition. Annu Rev Immunol. 2002;20:197–216. - PubMed
-
- Glaziou P, Floyd K, Raviglione M. Global burden and epidemiology of tuberculosis. Clin Chest Med. 2009;30:621–636, vii. - PubMed
-
- Miller EA, Ernst JD. Illuminating the black box of TNF action in tuberculous granulomas. Immunity. 2008;29:175–177. - PubMed
-
- Kaufmann SH. Tuberculosis: back on the immunologists' agenda. Immunity. 2006;24:351–357. - PubMed
-
- Keane J, Gershon S, Wise RP, Mirabile-Levens E, Kasznica J, et al. Tuberculosis associated with infliximab, a tumor necrosis factor alpha-neutralizing agent. N Engl J Med. 2001;345:1098–1104. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
