

Protein glutathionylation in the regulation of peroxiredoxins: a family of thiol-specific peroxidases that function as antioxidants, molecular chaperones, and signal modulators
- PMID: 22114845
- PMCID: PMC3270059
- DOI: 10.1089/ars.2011.4260
Protein glutathionylation in the regulation of peroxiredoxins: a family of thiol-specific peroxidases that function as antioxidants, molecular chaperones, and signal modulators
Abstract
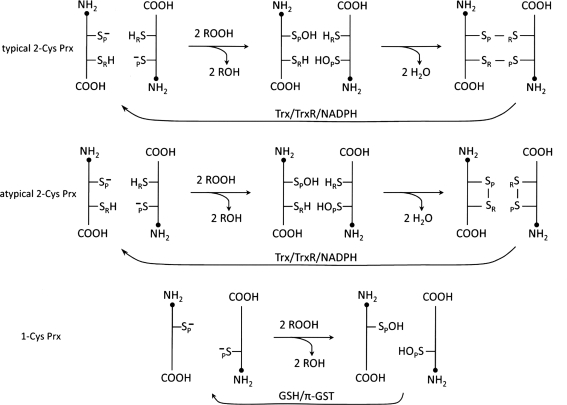
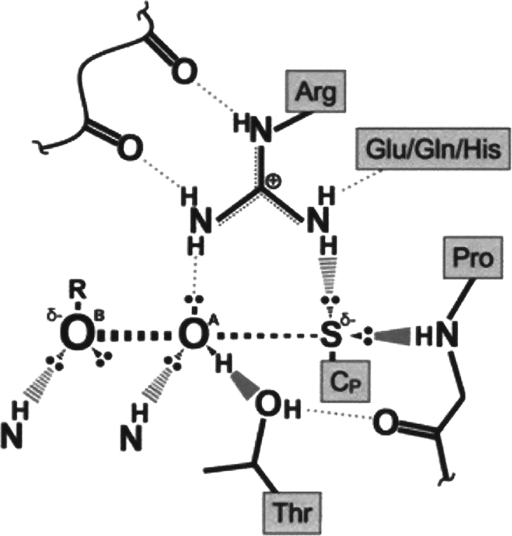
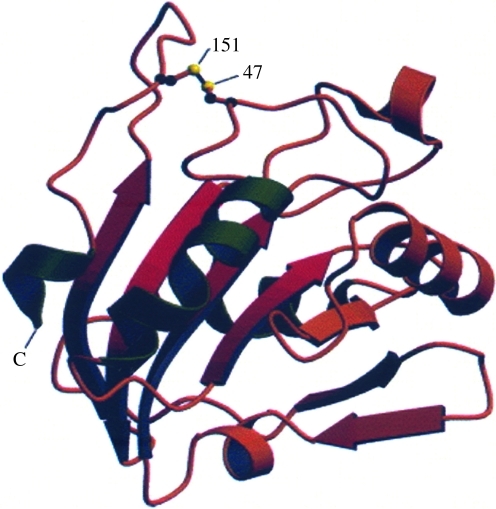
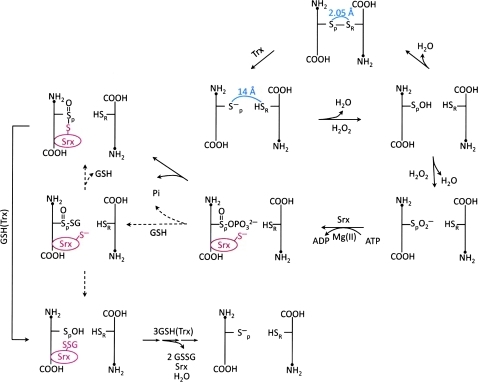
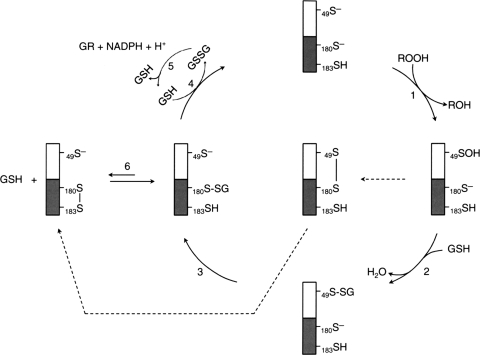
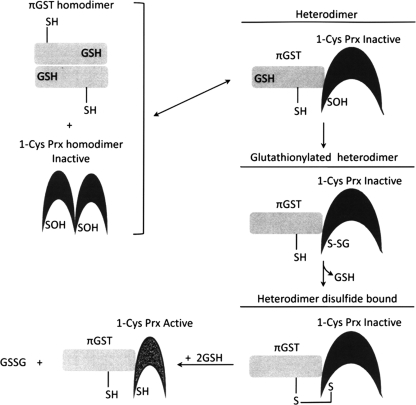
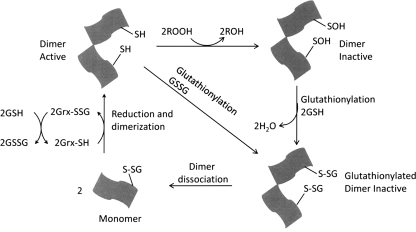
Significance: Reversible protein glutathionylation plays an important role in cellular regulation, signaling transduction, and antioxidant defense. This redox-sensitive mechanism is involved in regulating the functions of peroxiredoxins (Prxs), a family of ubiquitously expressed thiol-specific peroxidase enzymes. Glutathionylation of certain Prxs at their active-site cysteines not only provides reducing equivalents to support their peroxidase activity but also protects Prxs from irreversible hyperoxidation. Typical 2-Cys Prx also functions as a molecular chaperone when it exists as a decamer and/or higher molecular weight complexes. The hyperoxidized sulfinic derivative of 2-Cys Prx is reactivated by sulfiredoxin (Srx). In this review, the roles of glutathionylation in the regulation of Prxs are discussed with respect to their molecular structure and functions as antioxidants, molecular chaperones, and signal modulators.
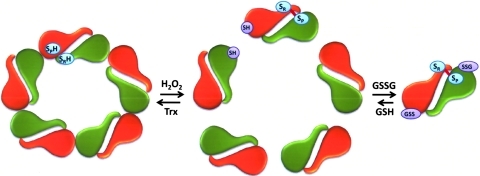
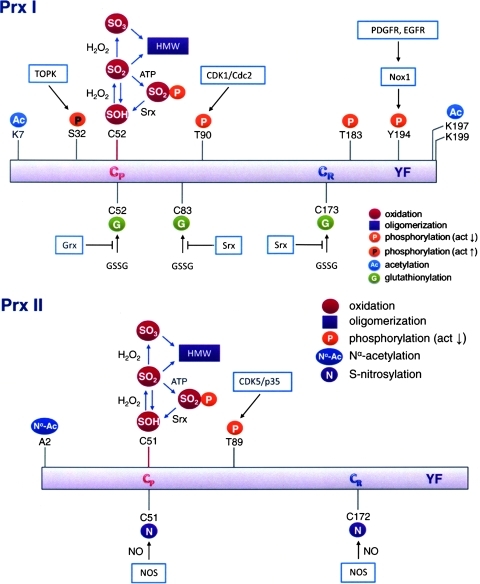
Recent advances: Recent findings reveal that glutathionylation regulates the quaternary structure of Prx. Glutathionylation of Prx I at Cys(83) converts the decameric Prx to its dimers with the loss of chaperone activity. The findings that dimer/oligomer structure specific Prx I binding proteins, e.g., phosphatase and tensin homolog (PTEN) and mammalian Ste20-like kinase-1 (MST1), regulate cell cycle and apoptosis, respectively, suggest a possible link between glutathionylation and those signaling pathways.
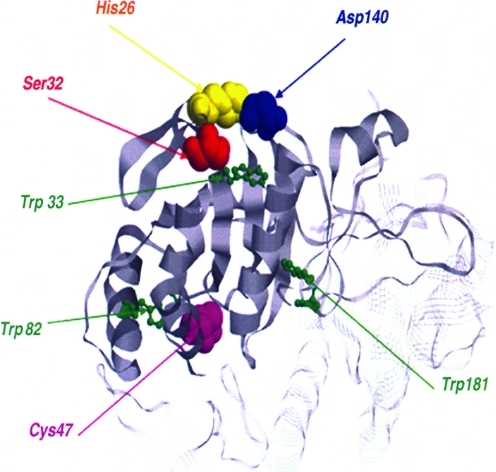
Critical issues: Knowing how glutathionylation affects the interaction between Prx I and its nearly 20 known interacting proteins, e.g., PTEN and MST1 kinase, would reveal new insights on the physiological functions of Prx.
Future directions: In vitro studies reveal that Prx oligomerization is linked to its functional changes. However, in vivo dynamics, including the effect by glutathionylation, and its physiological significance remain to be investigated.
Figures
References
-
- Adachi T. Pimentel DR. Heibeck T. Hou X. Lee YJ. Jiang B. Ido Y. Cohen RA. S-glutathiolation of Ras mediates redox-sensitive signaling by angiotensin II in vascular smooth muscle cells. J Biol Chem. 2004;279:29857–29862. - PubMed
-
- Alger HM. Williams DL. The disulfide redox system of Schistosoma mansoni and the importance of a multifunctional enzyme, thioredoxin glutathione reductase. Mol Biochem Parasitol. 2002;121:129–139. - PubMed
-
- Bae YS. Kang SW. Seo MS. Baines IC. Tekle E. Chock PB. Rhee SG. Epidermal growth factor (EGF)-induced generation of hydrogen peroxide. Role in EGF receptor-mediated tyrosine phosphorylation. J Biol Chem. 1997;272:217–221. - PubMed
-
- Barrett WC. DeGnore JP. Keng YF. Zhang ZY. Yim MB. Chock PB. Roles of superoxide radical anion in signal transduction mediated by reversible regulation of protein-tyrosine phosphatase 1B. J Biol Chem. 1999;274:34543–34546. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
