

Alternative splicing of the neurofibromatosis type I pre-mRNA
- PMID: 22115364
- PMCID: PMC3726530
- DOI: 10.1042/BSR20110060
Alternative splicing of the neurofibromatosis type I pre-mRNA
Abstract
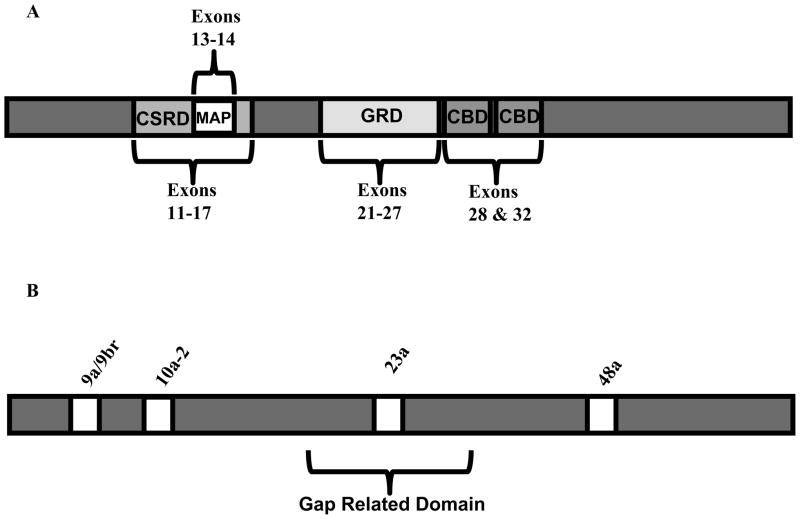
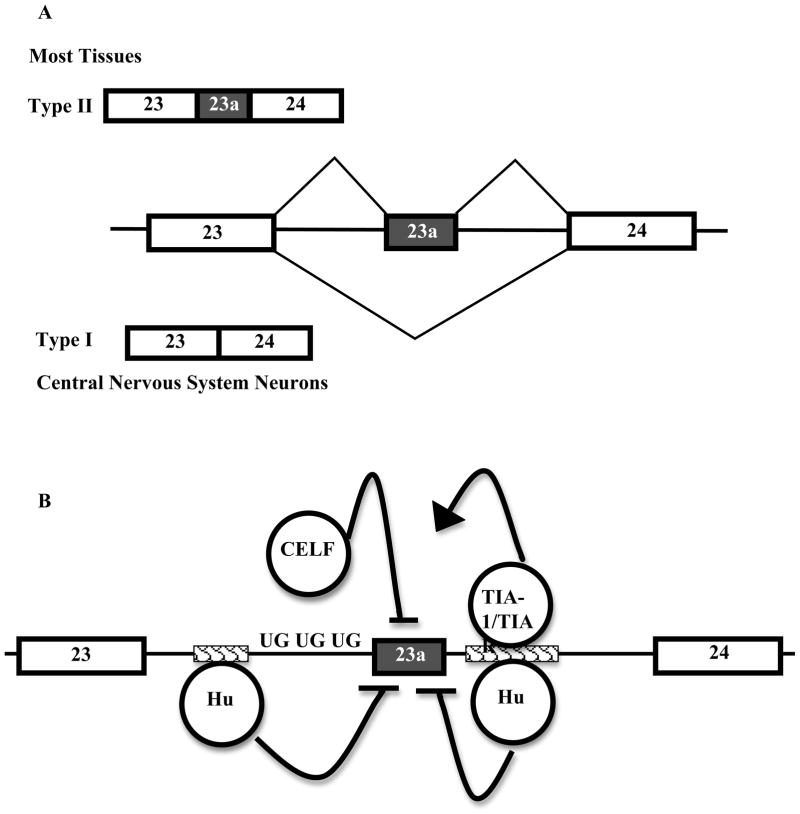
NF1 (neurofibromatosis type I) is a common genetic disease that affects one in 3500 individuals. The disease is completely penetrant but shows variable phenotypic expression in patients. NF1 is a large gene, and its pre-mRNA undergoes alternative splicing. The NF1 protein, neurofibromin, is involved in diverse signalling cascades. One of the best characterized functions of NF1 is its function as a Ras-GAP (GTPase-activating protein). NF1 exon 23a is an alternative exon that lies within the GAP-related domain of neurofibromin. This exon is predominantly included in most tissues, and it is skipped in CNS (central nervous system) neurons. The isoform in which exon 23a is skipped has 10 times higher Ras-GAP activity than the isoform in which exon 23a is included. Exon 23a inclusion is tightly regulated by at least three different families of RNA-binding proteins: CELF {CUG-BP (cytosine-uridine-guanine-binding protein) and ETR-3 [ELAV (embryonic lethal abnormal vision)-type RNA-binding protein]-like factor}, Hu and TIA-1 (T-cell intracellular antigen 1)/TIAR (T-cell intracellular antigen 1-related protein). The CELF and Hu proteins promote exon 23a skipping, while the TIA-1/TIAR proteins promote its inclusion. The widespread clinical variability that is observed among NF1 patients cannot be explained by NF1 mutations alone and it is believed that modifier genes may have a role in the variability. We suggest that the regulation of alternative splicing may act as a modifier to contribute to the variable expression in NF1 patients.
Figures
Similar articles
-
The neurofibromatosis type I pre-mRNA is a novel target of CELF protein-mediated splicing regulation.Nucleic Acids Res. 2010 Jan;38(1):253-64. doi: 10.1093/nar/gkp766. Epub 2009 Oct 23. Nucleic Acids Res. 2010. PMID: 19854948 Free PMC article.
-
Alternative splicing of the neurofibromatosis type 1 pre-mRNA is regulated by the muscleblind-like proteins and the CUG-BP and ELAV-like factors.BMC Mol Biol. 2012 Dec 10;13:35. doi: 10.1186/1471-2199-13-35. BMC Mol Biol. 2012. PMID: 23227900 Free PMC article.
-
Regulation of neuron-specific alternative splicing of neurofibromatosis type 1 pre-mRNA.Mol Cell Biol. 2008 Feb;28(4):1240-51. doi: 10.1128/MCB.01509-07. Epub 2007 Dec 17. Mol Cell Biol. 2008. PMID: 18086893 Free PMC article.
-
Alternative Splicing of Exon 23a in Neurofibromatosis Type 1 Pre-mRNA: Its Contribution to the Protein Structure and Function of Neurofibromin.Wiley Interdiscip Rev RNA. 2025 Jul-Aug;16(4):e70021. doi: 10.1002/wrna.70021. Wiley Interdiscip Rev RNA. 2025. PMID: 40812791 Free PMC article. Review.
-
Neurofibromatosis 1: closing the GAP between mice and men.Curr Opin Genet Dev. 2003 Feb;13(1):20-7. doi: 10.1016/s0959-437x(02)00015-1. Curr Opin Genet Dev. 2003. PMID: 12573431 Review.
Cited by
-
T-Cell Intracellular Antigen 1-Like Protein in Physiology and Pathology.Int J Mol Sci. 2022 Jul 16;23(14):7836. doi: 10.3390/ijms23147836. Int J Mol Sci. 2022. PMID: 35887183 Free PMC article. Review.
-
Oncogenic alternative splicing switches: role in cancer progression and prospects for therapy.Int J Cell Biol. 2013;2013:962038. doi: 10.1155/2013/962038. Epub 2013 Oct 27. Int J Cell Biol. 2013. PMID: 24285959 Free PMC article. Review.
-
Isoform-specific NF1 mRNA levels correlate with disease severity in Neurofibromatosis type 1.Orphanet J Rare Dis. 2019 Nov 15;14(1):261. doi: 10.1186/s13023-019-1223-1. Orphanet J Rare Dis. 2019. PMID: 31730495 Free PMC article.
-
Neurofibromatosis type 1 alternative splicing is a key regulator of Ras signaling in neurons.Mol Cell Biol. 2014 Jun;34(12):2188-97. doi: 10.1128/MCB.00019-14. Epub 2014 Apr 7. Mol Cell Biol. 2014. PMID: 24710274 Free PMC article.
-
Immortalization of human normal and NF1 neurofibroma Schwann cells.Lab Invest. 2016 Oct;96(10):1105-15. doi: 10.1038/labinvest.2016.88. Epub 2016 Sep 12. Lab Invest. 2016. PMID: 27617404
References
-
- Pan Q, Shai O, Lee LJ, Frey BJ, Blencowe BJ. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat Genet. 2008;40:1413–1415. - PubMed
-
- Black DL. Mechanisms of alternative pre-messenger RNA splicing. Annu Rev Biochem. 2003;72:291–336. - PubMed
-
- Black DL, Grabowski PJ. Alternative pre-mRNA splicing and neuronal function. Prog Mol Subcell Biol. 2003;31:187–216. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous