

Rapid reversal of impaired inhibitory and excitatory transmission but not spine dysgenesis in a mouse model of mental retardation
- PMID: 22124149
- PMCID: PMC3381309
- DOI: 10.1113/jphysiol.2011.219907
Rapid reversal of impaired inhibitory and excitatory transmission but not spine dysgenesis in a mouse model of mental retardation
Abstract
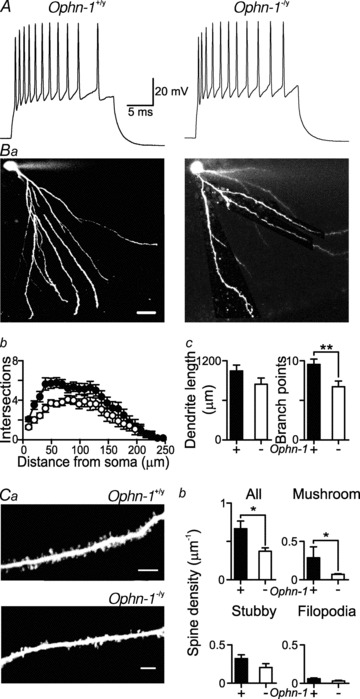
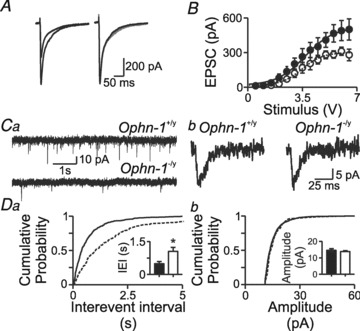
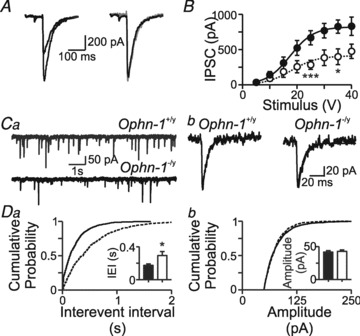
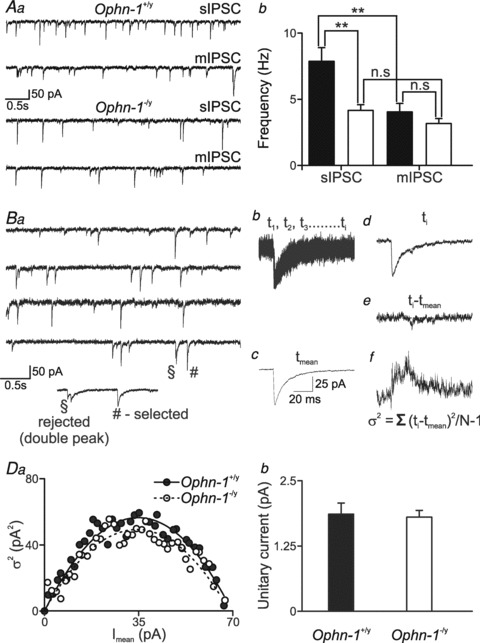
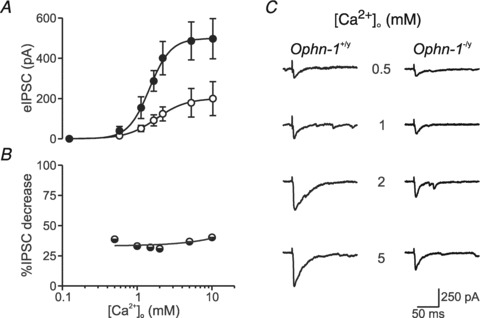
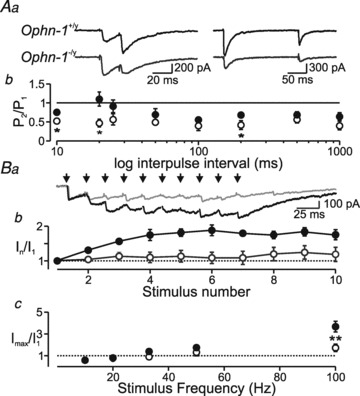
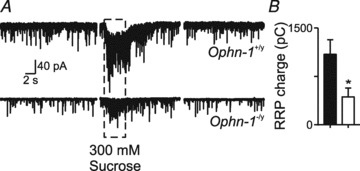
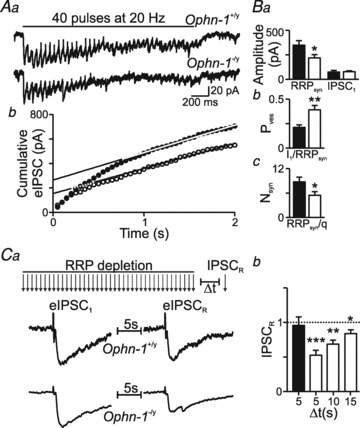
Intellectual disability affects 2-3% of the population: those due to mutations of the X-chromosome are a major cause of moderate to severe cases (1.8/1000 males). Established theories ascribe the cellular aetiology of intellectual disability to malformations of dendritic spines. Recent work has identified changes in synaptic physiology in some experimental models. Here, we investigated the pathophysiology of a mouse model of intellectual disability using electrophysiological recordings combined with confocal imaging of dentate gyrus granule neurons. Lack of oligophrenin-1 resulted in reductions in dendritic tree complexity and mature dendritic spine density and in evoked and spontaneous EPSCs and IPSCs. In the case of inhibitory transmission, the physiological change was associated with a reduction in the readily releasable pool and vesicle recycling which impaired the efficiency of inhibitory synaptic transmission. Acute inhibition of the downstream signalling pathway of oligophrenin-1 fully reversed the functional changes in synaptic transmission but not the dendritic abnormalities. The impaired inhibitory (as well as excitatory) synaptic transmission at frequencies associated with cognitive function suggests a cellular mechanism for the intellectual disability, because cortical oscillations associated with cognition normally depend on inhibitory neurons firing on every cycle.
Figures
Comment in
-
Synaptic activity in X-linked mental retardation: a thorny issue.J Physiol. 2012 Feb 15;590(4):653. doi: 10.1113/jphysiol.2011.226019. J Physiol. 2012. PMID: 22344981 Free PMC article. No abstract available.
References
-
- Ashton AC, Ushkaryov YA. Properties of synaptic vesicle pools in mature central nerve terminals. J Biol Chem. 2005;280:37278–37288. - PubMed
-
- Bartos M, Vida I, Jonas P. Synaptic mechanisms of synchronized gamma oscillations in inhibitory interneuron networks. Nat Rev Neurosci. 2007;8:45–56. - PubMed
-
- Billuart P, Bienvenu T, Ronce N, des Portes V, Vinet MC, Zemni R, Roest CH, Carrie A, Fauchereau F, Cherry M, Briault S, Hamel B, Fryns JP, Beldjord C, Kahn A, Moraine C, Chelly J. Oligophrenin-1 encodes a RhoGAP protein involved in X-linked mental retardation. Nature. 1998;392:923–926. - PubMed
-
- Chelly J, Khelfaoui M, Francis F, Cherif B, Bienvenu T. Genetics and pathophysiology of mental retardation. Eur J Hum Genet. 2006;14:701–713. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases