

Generation and replication-dependent dilution of 5fC and 5caC during mouse preimplantation development
- PMID: 22124233
- PMCID: PMC3357997
- DOI: 10.1038/cr.2011.189
Generation and replication-dependent dilution of 5fC and 5caC during mouse preimplantation development
Abstract
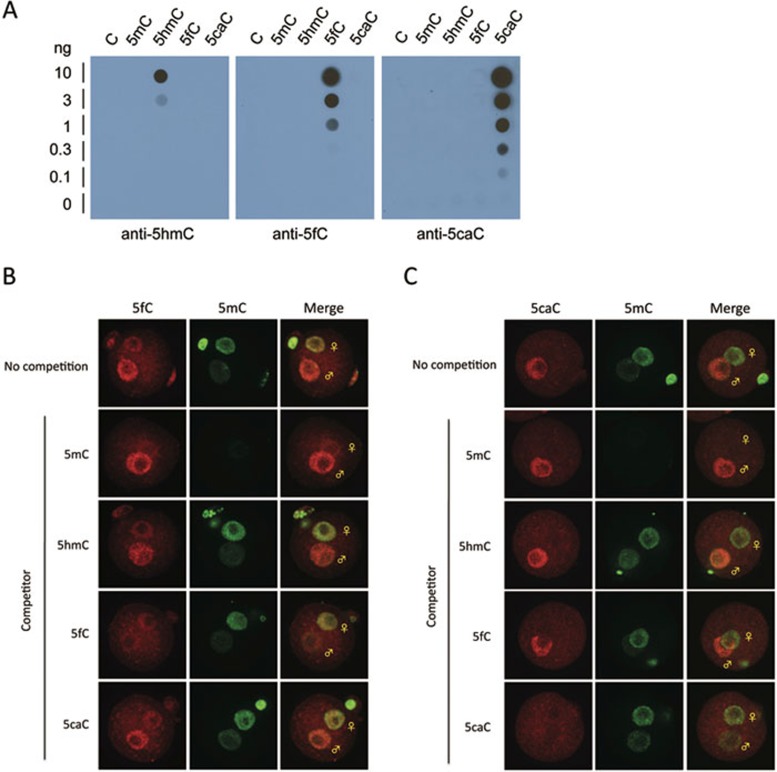
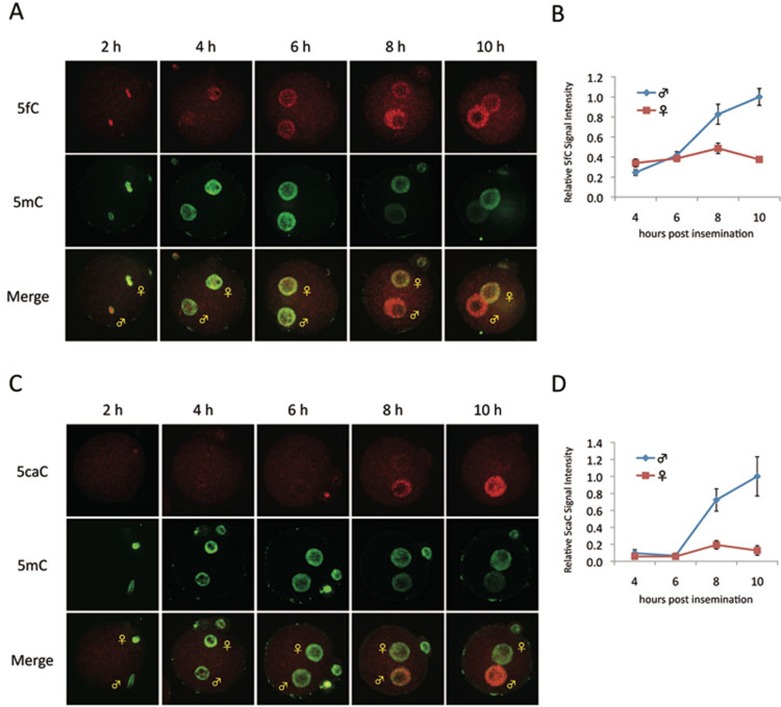
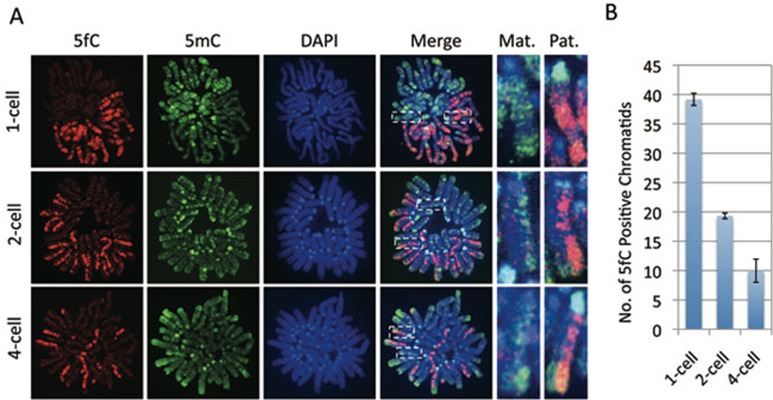
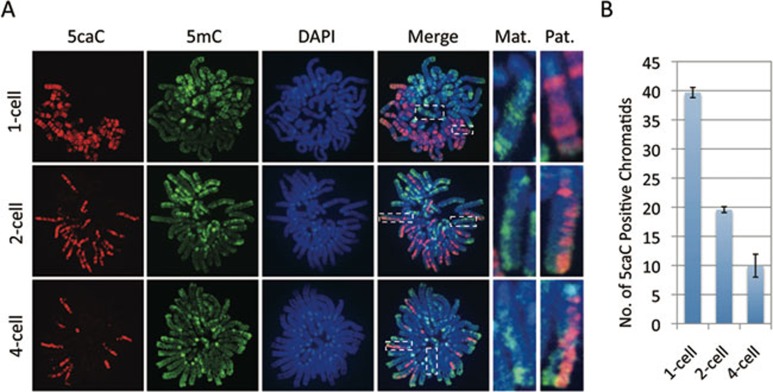
One of the recent advances in the epigenetic field is the demonstration that the Tet family of proteins are capable of catalyzing conversion of 5-methylcytosine (5mC) of DNA to 5-hydroxymethylcytosine (5hmC). Interestingly, recent studies have shown that 5hmC can be further oxidized by Tet proteins to generate 5-formylcytosine (5fC) and 5-carboxylcytosine (5caC), which can be removed by thymine DNA glycosylase (TDG). To determine whether Tet-catalyzed conversion of 5mC to 5fC and 5caC occurs in vivo in zygotes, we generated antibodies specific for 5fC and 5caC. By immunostaining, we demonstrate that loss of 5mC in the paternal pronucleus is concurrent with the appearance of 5fC and 5caC, similar to that of 5hmC. Importantly, instead of being quickly removed through an enzyme-catalyzed process, both 5fC and 5caC exhibit replication-dependent dilution during mouse preimplantation development. These results not only demonstrate the conversion of 5mC to 5fC and 5caC in zygotes, but also indicate that both 5fC and 5caC are relatively stable and may be functional during preimplantation development. Together with previous studies, our study suggests that Tet-catalyzed conversion of 5mC to 5hmC/5fC/5caC followed by replication-dependent dilution accounts for paternal DNA demethylation during preimplantation development.
Figures
Comment in
-
Active DNA demethylation by oxidation and repair.Cell Res. 2011 Dec;21(12):1649-51. doi: 10.1038/cr.2011.140. Epub 2011 Aug 23. Cell Res. 2011. PMID: 21862972 Free PMC article. No abstract available.
References
-
- Cedar H, Bergman Y. Linking DNA methylation and histone modification: patterns and paradigms. Nat Rev Genet. 2009;10:295–304. - PubMed
-
- Sasaki H, Matsui Y. Epigenetic events in mammalian germ-cell development: reprogramming and beyond. Nat Rev Genet. 2008;9:129–140. - PubMed
-
- Suzuki MM, Bird A. DNA methylation landscapes: provocative insights from epigenomics. Nat Rev Genet. 2008;9:465–476. - PubMed
-
- Goll MG, Bestor TH. Eukaryotic cytosine methyltransferases. Annu Rev Biochem. 2005;74:481–514. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
