

Physiological IRE-1-XBP-1 and PEK-1 signaling in Caenorhabditis elegans larval development and immunity
- PMID: 22125500
- PMCID: PMC3219621
- DOI: 10.1371/journal.pgen.1002391
Physiological IRE-1-XBP-1 and PEK-1 signaling in Caenorhabditis elegans larval development and immunity
Abstract
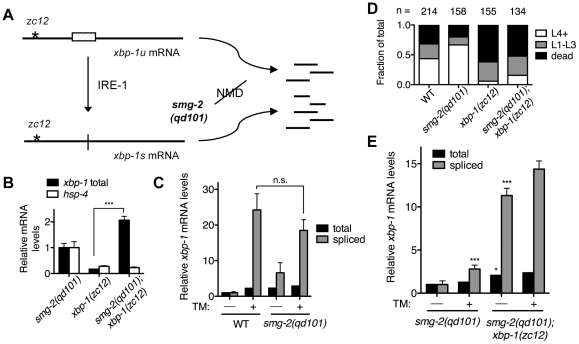
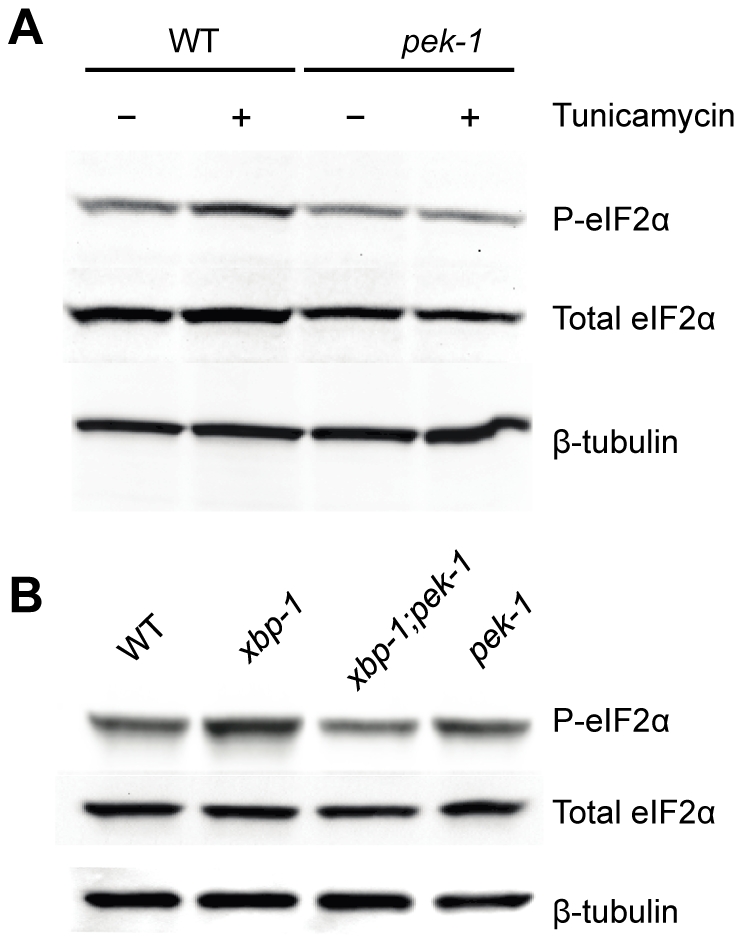
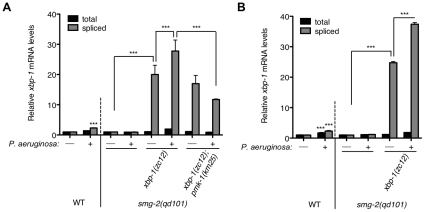
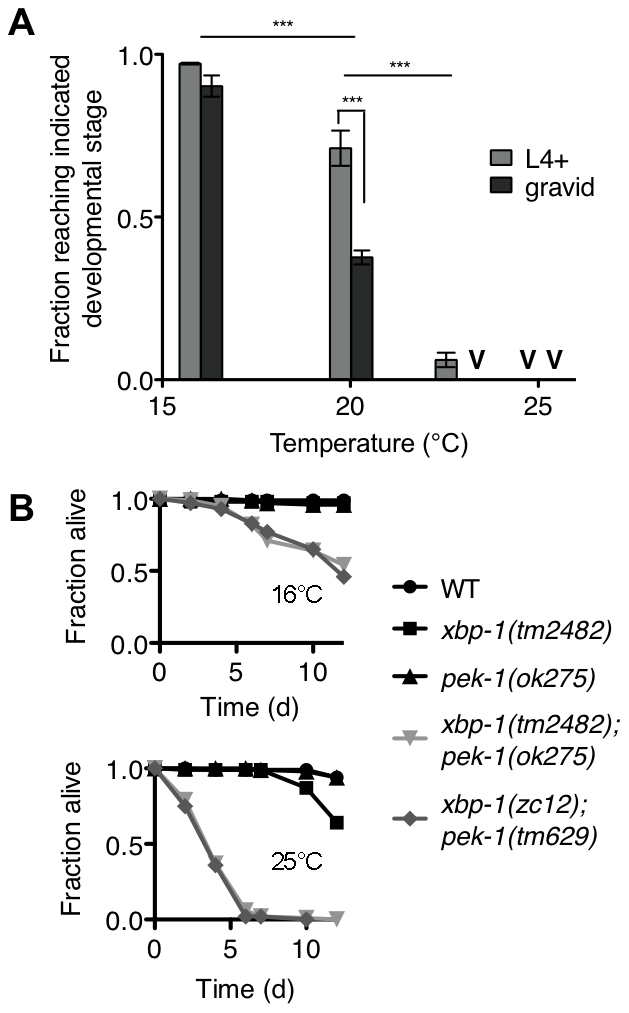
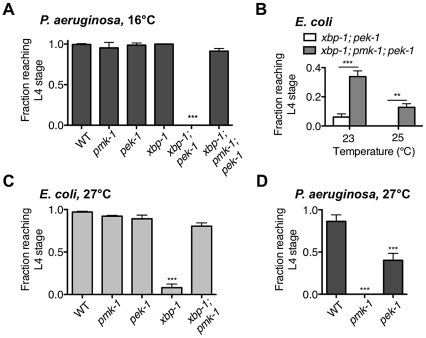
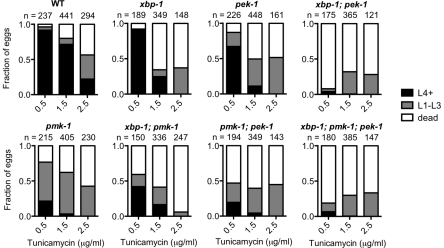
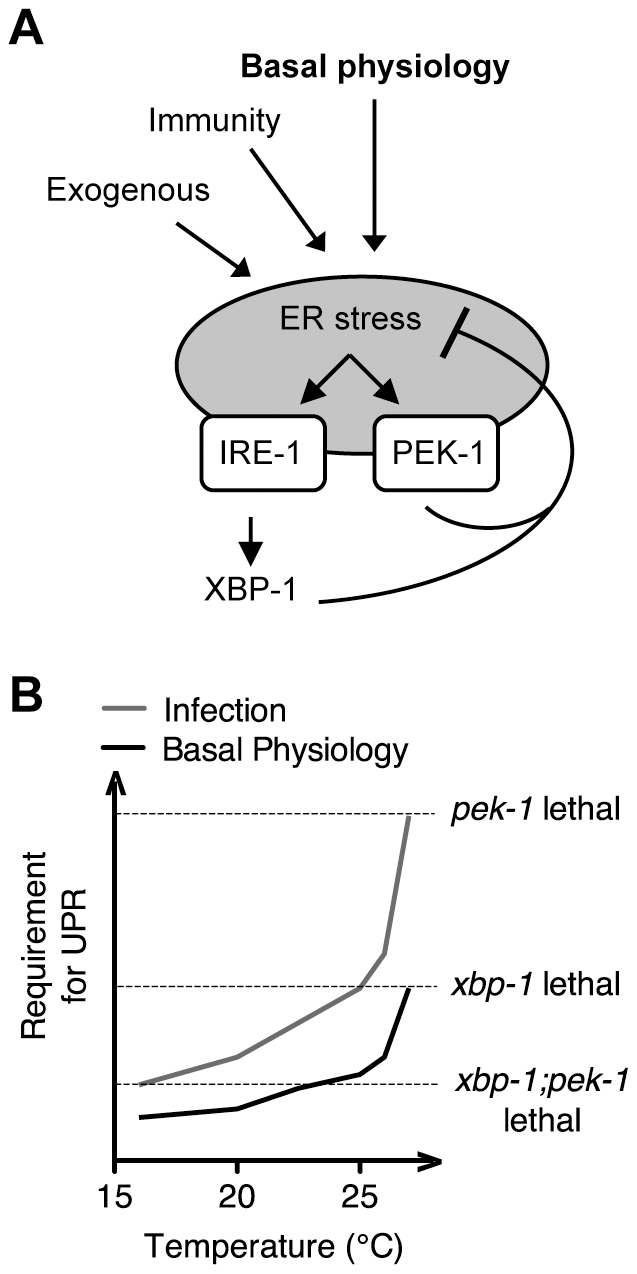
Endoplasmic reticulum (ER) stress activates the Unfolded Protein Response, a compensatory signaling response that is mediated by the IRE-1, PERK/PEK-1, and ATF-6 pathways in metazoans. Genetic studies have implicated roles for UPR signaling in animal development and disease, but the function of the UPR under physiological conditions, in the absence of chemical agents administered to induce ER stress, is not well understood. Here, we show that in Caenorhabditis elegans XBP-1 deficiency results in constitutive ER stress, reflected by increased basal levels of IRE-1 and PEK-1 activity under physiological conditions. We define a dynamic, temperature-dependent requirement for XBP-1 and PEK-1 activities that increases with immune activation and at elevated physiological temperatures in C. elegans. Our data suggest that the negative feedback loops involving the activation of IRE-1-XBP-1 and PEK-1 pathways serve essential roles, not only at the extremes of ER stress, but also in the maintenance of ER homeostasis under physiological conditions.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Mori K. Signaling pathways in the unfolded protein response: development from yeast to mammals. Journal of Biochemistry. 2009;146:743–750. - PubMed
-
- Ron D, Walter P. Signal integration in the endoplasmic reticulum unfolded protein response. Nature Reviews Molecular Cell Biology. 2007;8:519–529. - PubMed
-
- Schröder M, Kaufman RJ. The mammalian unfolded protein response. Annual Reviews in Biochemistry. 2005;74:739–789. - PubMed
-
- Calfon M, Zeng HQ, Urano F, Till JH, Hubbard SR, et al. IRE1 couples endoplasmic reticulum load to secretory capacity by processing the XBP-1 mRNA. Nature. 2002;415:92–96. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
