

Eco-evolutionary feedback and the invasion of cooperation in prisoner's dilemma games
- PMID: 22125615
- PMCID: PMC3220694
- DOI: 10.1371/journal.pone.0027523
Eco-evolutionary feedback and the invasion of cooperation in prisoner's dilemma games
Abstract
Unveiling the origin and forms of cooperation in nature poses profound challenges in evolutionary ecology. The prisoner's dilemma game is an important metaphor for studying the evolution of cooperation. We here classified potential mechanisms for cooperation evolution into schemes of frequency- and density-dependent selection, and focused on the density-dependent selection in the ecological prisoner's dilemma games. We found that, although assortative encounter is still the necessary condition in ecological games for cooperation evolution, a harsh environment, indicated by a high mortality, can foster the invasion of cooperation. The Hamilton rule provides a fundamental condition for the evolution of cooperation by ensuring an enhanced relatedness between players in low-density populations. Incorporating ecological dynamics into evolutionary games opens up a much wider window for the evolution of cooperation, and exhibits a variety of complex behaviors of dynamics, such as limit and heteroclinic cycles. An alternative evolutionary, or rather succession, sequence was proposed that cooperation first appears in harsh environments, followed by the invasion of defection, which leads to a common catastrophe. The rise of cooperation (and altruism), thus, could be much easier in the density-dependent ecological games than in the classic frequency-dependent evolutionary games.
Conflict of interest statement
Figures
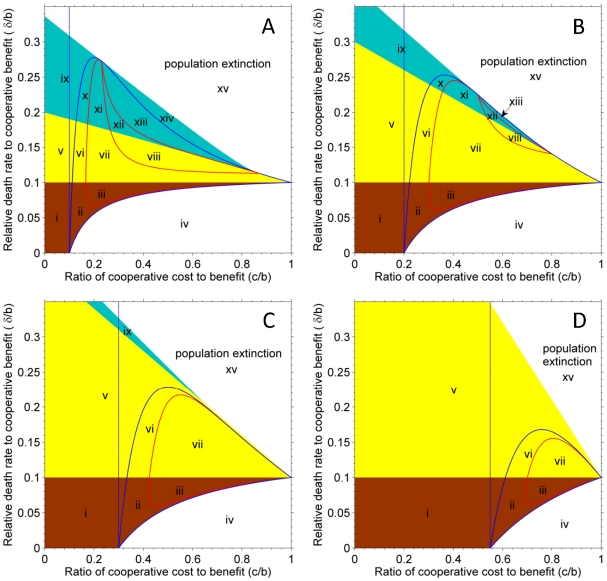
 , have two boundary equilibriums: one for cooperators; the other for defectors. Yellow part,
, have two boundary equilibriums: one for cooperators; the other for defectors. Yellow part,  , has only one boundary equilibrium for cooperators. Cyan part,
, has only one boundary equilibrium for cooperators. Cyan part,  , has two boundary equilibriums both for cooperators. The area encircled by blue curves indicates the existence of interior equilibrium of cooperation-defect coexistence. The three red lines on (A) and (B), from bottom to top, indicate the node-focus bifurcation, Hopf bifurcation and the heteroclinic bifurcation, respectively, and the red lines on (C) and (D) indicate the node-focus bifurcation. Area on the left side of the vertical dotted line indicates the invasion condition for cooperation in the frequency-dependent selection (i.e.
, has two boundary equilibriums both for cooperators. The area encircled by blue curves indicates the existence of interior equilibrium of cooperation-defect coexistence. The three red lines on (A) and (B), from bottom to top, indicate the node-focus bifurcation, Hopf bifurcation and the heteroclinic bifurcation, respectively, and the red lines on (C) and (D) indicate the node-focus bifurcation. Area on the left side of the vertical dotted line indicates the invasion condition for cooperation in the frequency-dependent selection (i.e.  ). Parameters are
). Parameters are  and m = 0.1, 0.2, 0.3 and 0.55, respectively, for panel (A) to (D).
and m = 0.1, 0.2, 0.3 and 0.55, respectively, for panel (A) to (D).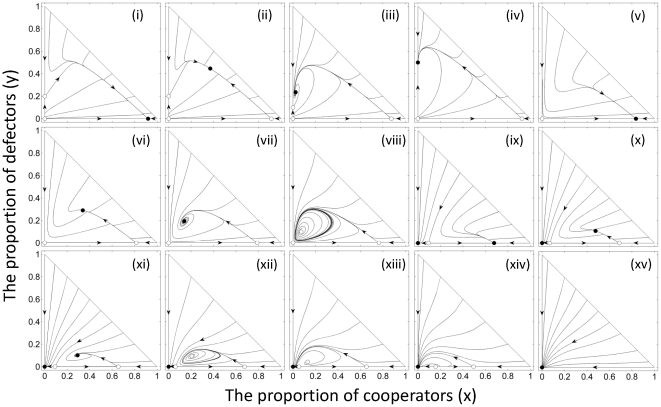
 ,
,  ,
,  for all diagrams, except
for all diagrams, except  and
and  for (i),
for (i),  and
and  for (ii),
for (ii),  and
and  for (iii),
for (iii),  and
and  for (iv),
for (iv),  and
and  for (v),
for (v),  and
and  for (vi),
for (vi),  and
and  for (vii),
for (vii),  and
and  for (viii),
for (viii),  and
and  for (ix),
for (ix),  and
and  for (x),
for (x),  and
and  for (xi),
for (xi),  and
and  for (xii),
for (xii),  and
and  for (xiii),
for (xiii),  and
and  for (xiv), and
for (xiv), and  and
and  for (xv).
for (xv).
References
-
- Wilkinson GS. Reciprocal food-sharing in the vampire bat. Nature. 1984;308:181–184.
-
- Sella G. Reciprocal egg trading and brood care in a hermaphroditic polychaete worm. Animal Behaviour. 1985;33:938–944.
-
- Pierce NE, Kitching RL, Buckley RC, Taylor MF, Benbow KF. The costs and benefits of cooperation between the Australian Lycaenid butterfly, Jalmenus evagoras, and its attendant ants. Behavioral Ecology and Sociobiology. 1987;21:237–248.
-
- Hemelrijk CK. Models of, and tests for, reciprocity, unidirectionality and other social interaction patterns at a group level. Animal Behaviour. 1990;39:1013–1029.
-
- Lombardo MP. Tree swallows and Tit for Tat: response to koening. Ethol Sociobiol. 1990;11:521–528.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
