

Role of JNK in mammary gland development and breast cancer
- PMID: 22127926
- PMCID: PMC3261359
- DOI: 10.1158/0008-5472.CAN-11-1628
Role of JNK in mammary gland development and breast cancer
Abstract
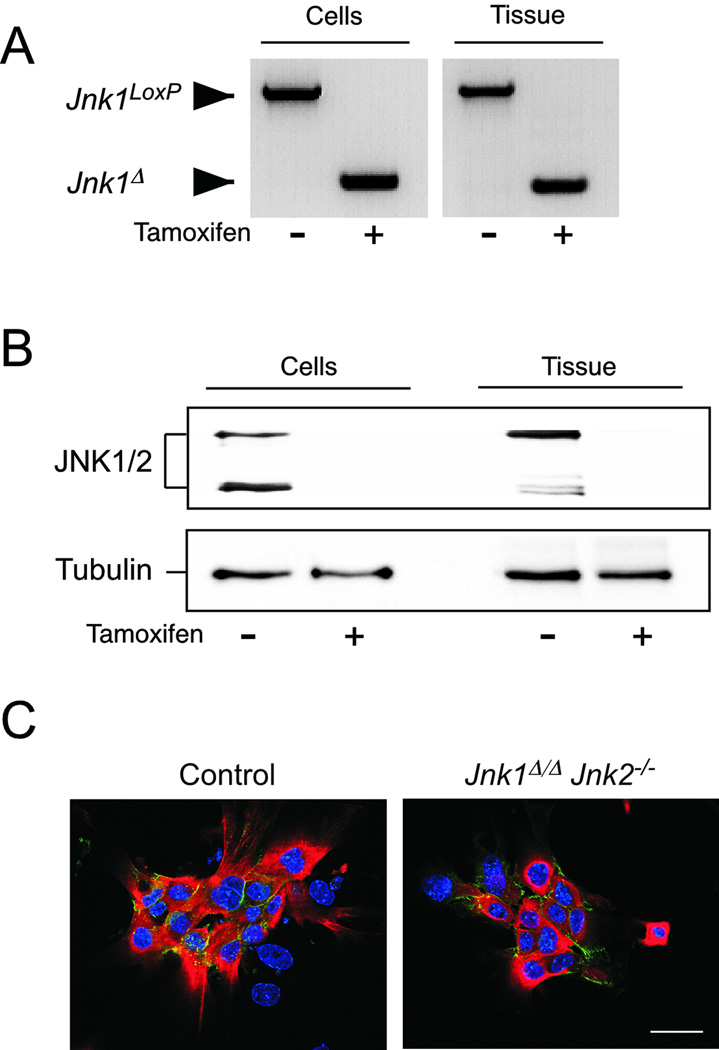
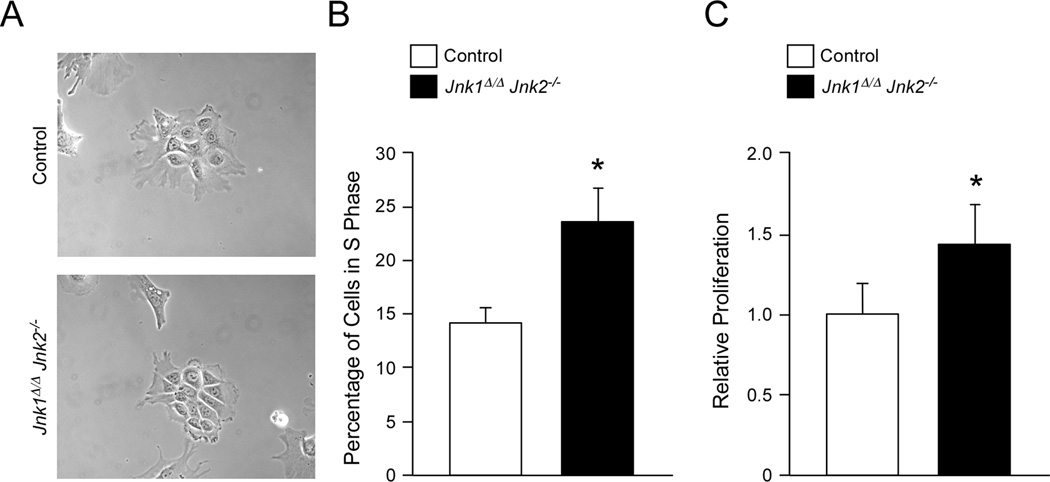
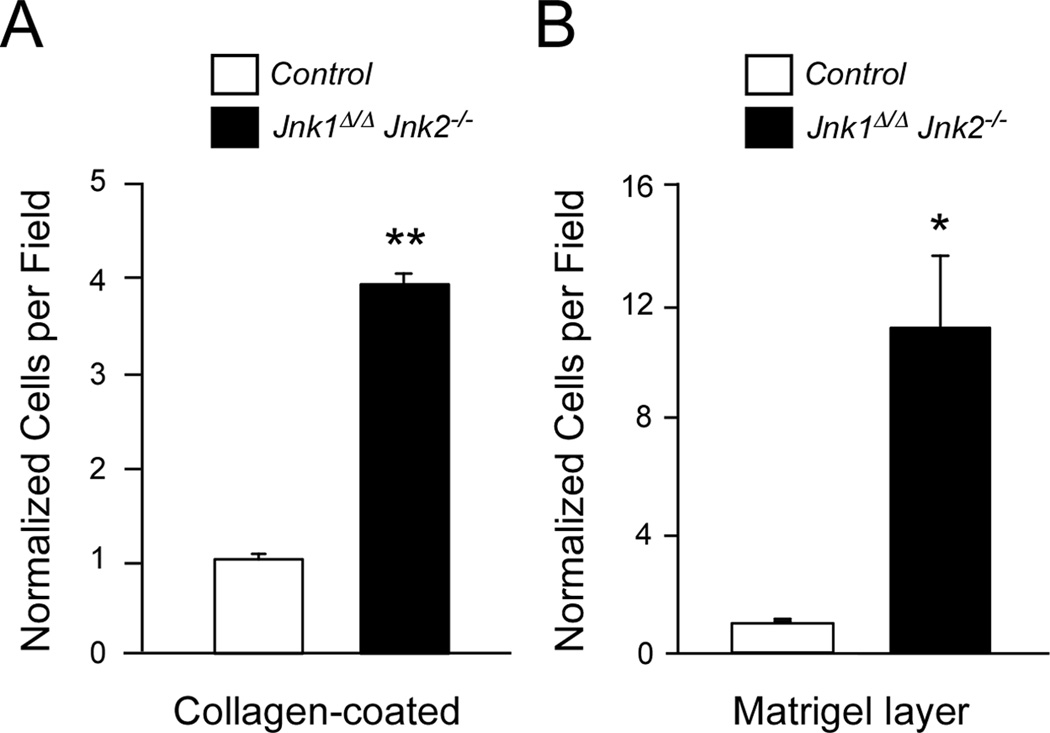
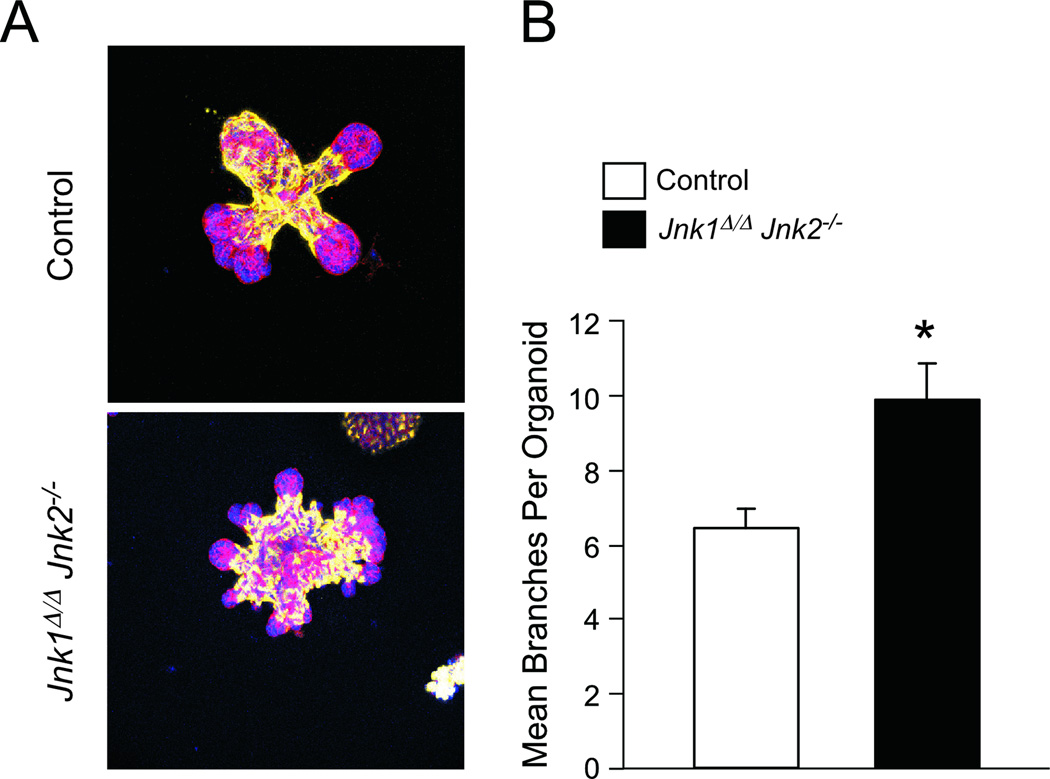
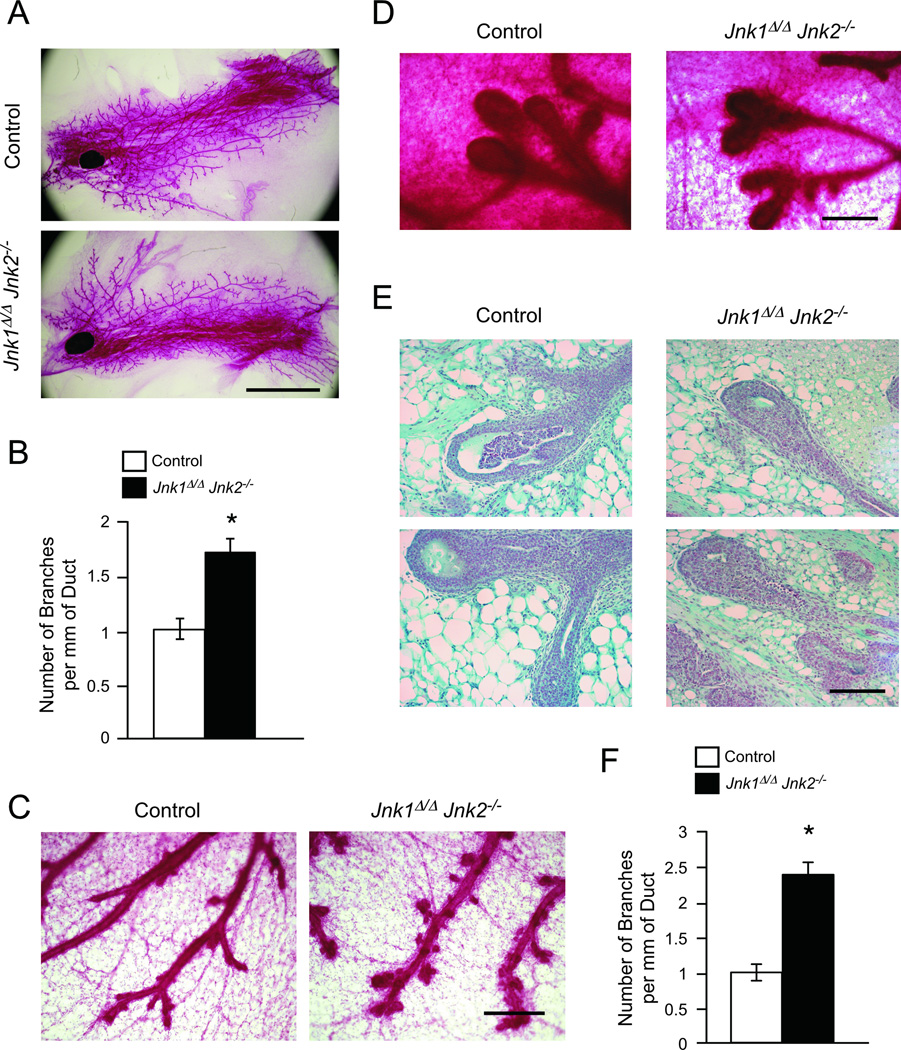
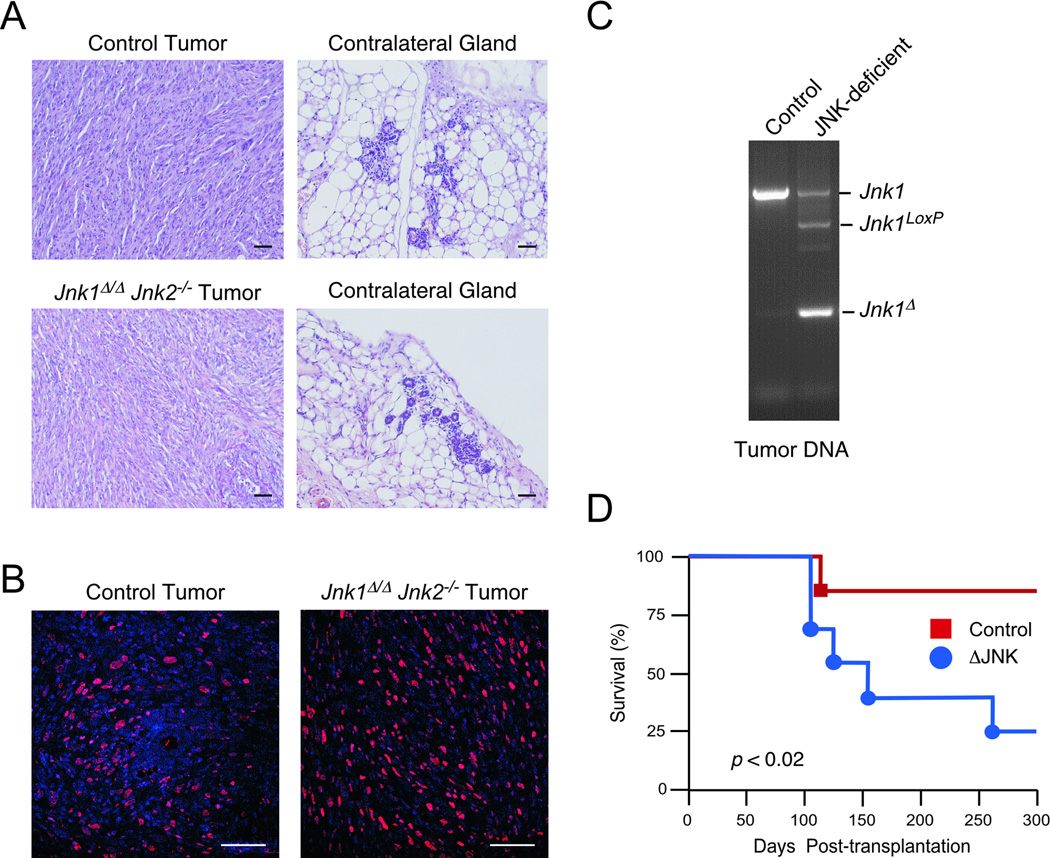
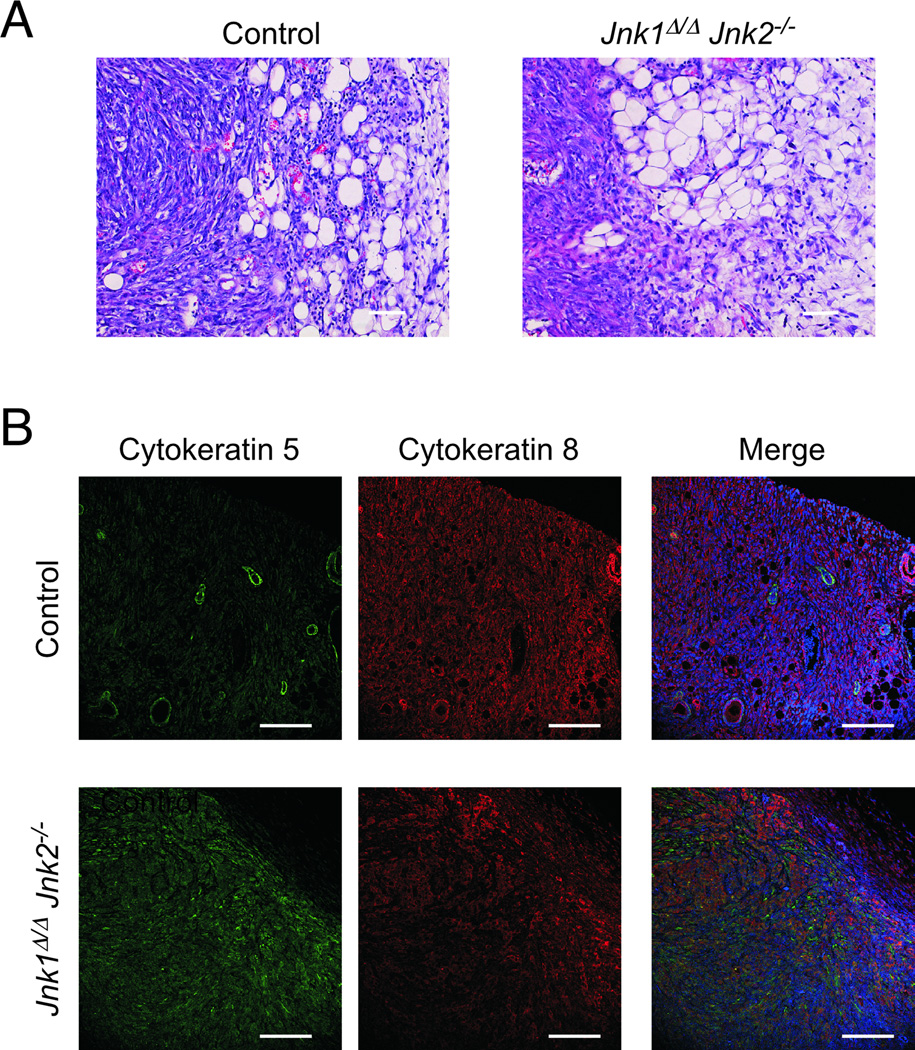
cJun NH(2)-terminal kinase (JNK) signaling has been implicated in the developmental morphogenesis of epithelial organs. In this study, we employed a compound deletion of the murine Jnk1 and Jnk2 genes in the mammary gland to evaluate the requirement for these ubiquitously expressed genes in breast development and tumorigenesis. JNK1/2 was not required for breast epithelial cell proliferation or motility. However, JNK1/2 deficiency caused increased branching morphogenesis and defects in the clearance of lumenal epithelial cells. In the setting of breast cancer development, JNK1/2 deficiency significantly increased tumor formation. Together, these findings established that JNK signaling is required for normal mammary gland development and that it has a suppressive role in mammary tumorigenesis.
Conflict of interest statement
No potential conflicts of interest were disclosed.
Figures
Similar articles
-
Role of JNK in a Trp53-dependent mouse model of breast cancer.PLoS One. 2010 Aug 30;5(8):e12469. doi: 10.1371/journal.pone.0012469. PLoS One. 2010. PMID: 20814571 Free PMC article.
-
Differential transmission of MEKK1 morphogenetic signals by JNK1 and JNK2.Development. 2008 Jan;135(1):23-32. doi: 10.1242/dev.007120. Epub 2007 Nov 21. Development. 2008. PMID: 18032450
-
Combined Activities of JNK1 and JNK2 in Hepatocytes Protect Against Toxic Liver Injury.Gastroenterology. 2016 Apr;150(4):968-81. doi: 10.1053/j.gastro.2015.12.019. Epub 2015 Dec 19. Gastroenterology. 2016. PMID: 26708719 Free PMC article.
-
Key signalling nodes in mammary gland development and cancer. Mitogen-activated protein kinase signalling in experimental models of breast cancer progression and in mammary gland development.Breast Cancer Res. 2009;11(5):209. doi: 10.1186/bcr2361. Breast Cancer Res. 2009. PMID: 19818165 Free PMC article. Review.
-
cJun NH2-terminal kinase 1 (JNK1): roles in metabolic regulation of insulin resistance.Trends Biochem Sci. 2010 Sep;35(9):490-6. doi: 10.1016/j.tibs.2010.04.004. Epub 2010 May 7. Trends Biochem Sci. 2010. PMID: 20452774 Free PMC article. Review.
Cited by
-
The role of MAPK signaling pathway in the Her-2-positive meningiomas.Oncol Rep. 2016 Aug;36(2):685-95. doi: 10.3892/or.2016.4849. Epub 2016 Jun 2. Oncol Rep. 2016. PMID: 27279438 Free PMC article.
-
JNK2 Is Required for the Tumorigenic Properties of Melanoma Cells.ACS Chem Biol. 2019 Jul 19;14(7):1426-1435. doi: 10.1021/acschembio.9b00083. Epub 2019 May 17. ACS Chem Biol. 2019. PMID: 31063355 Free PMC article.
-
JNK Promotes Epithelial Cell Anoikis by Transcriptional and Post-translational Regulation of BH3-Only Proteins.Cell Rep. 2017 Nov 14;21(7):1910-1921. doi: 10.1016/j.celrep.2017.10.067. Cell Rep. 2017. PMID: 29141222 Free PMC article.
-
JNK in Tumor Microenvironment: Present Findings and Challenges in Clinical Translation.Cancers (Basel). 2021 May 3;13(9):2196. doi: 10.3390/cancers13092196. Cancers (Basel). 2021. PMID: 34063627 Free PMC article. Review.
-
c-Jun N-terminal kinase 2 prevents luminal cell commitment in normal mammary glands and tumors by inhibiting p53/Notch1 and breast cancer gene 1 expression.Oncotarget. 2015 May 20;6(14):11863-81. doi: 10.18632/oncotarget.3787. Oncotarget. 2015. PMID: 25970777 Free PMC article.
References
-
- Davis RJ. Signal transduction by the JNK group of MAP kinases. Cell. 2000;103:239–252. - PubMed
-
- Xia Y, Karin M. The control of cell motility and epithelial morphogenesis by Jun kinases. Trends Cell Biol. 2004;14:94–101. - PubMed
-
- Ip YT, Davis RJ. Signal transduction by the c-Jun N-terminal kinase (JNK)--from inflammation to development. Curr Opin Cell Biol. 1998;10:205–219. - PubMed
-
- Zeitlinger J, Bohmann D. Thorax closure in Drosophila: involvement of Fos and the JNK pathway. Development. 1999;126:3947–3956. - PubMed
-
- Agnes F, Suzanne M, Noselli S. The Drosophila JNK pathway controls the morphogenesis of imaginal discs during metamorphosis. Development. 1999;126:5453–5462. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
