

Integrin inside-out signaling and the immunological synapse
- PMID: 22129583
- PMCID: PMC3294052
- DOI: 10.1016/j.ceb.2011.10.004
Integrin inside-out signaling and the immunological synapse
Abstract
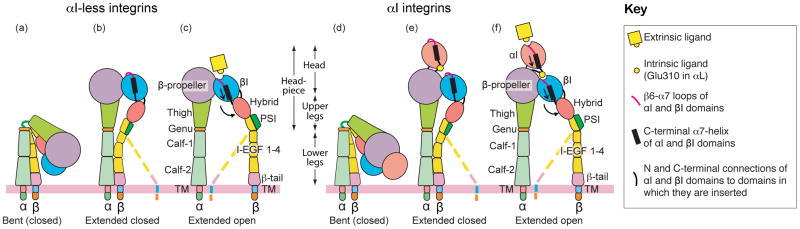
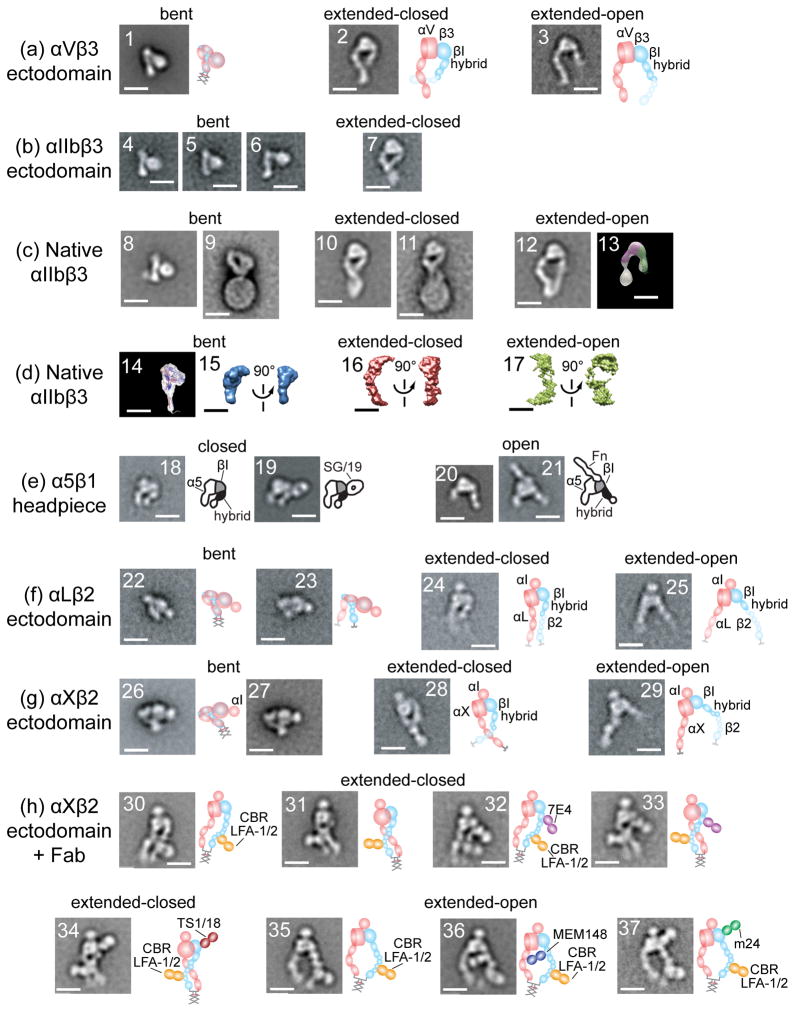
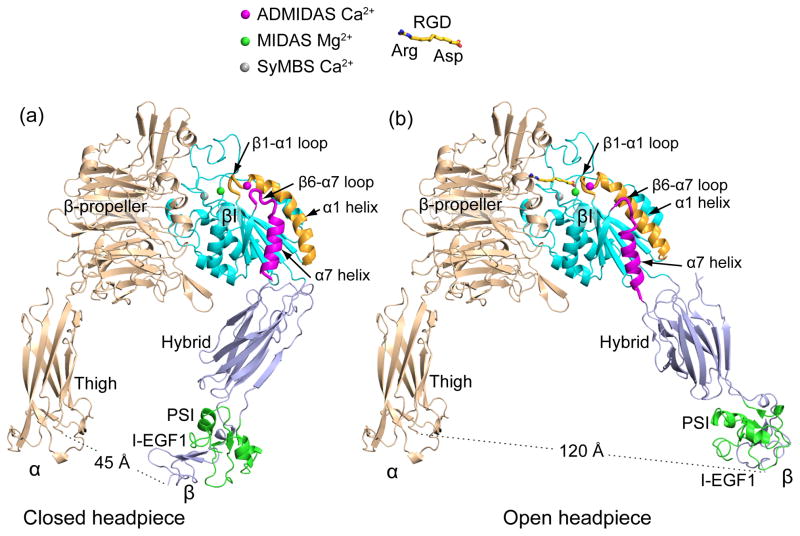
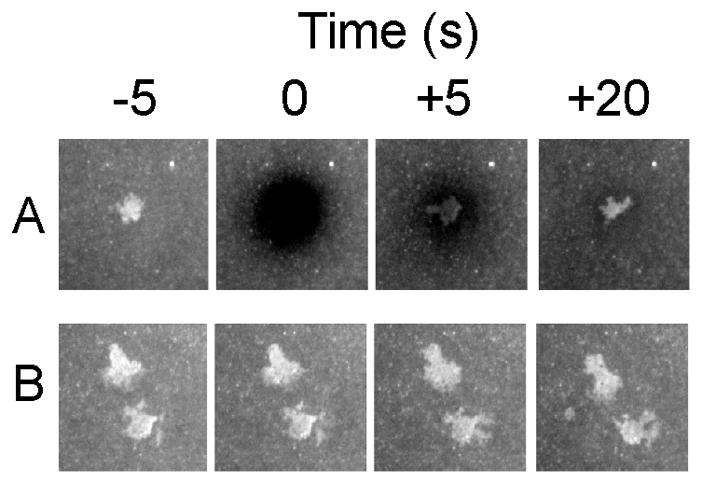
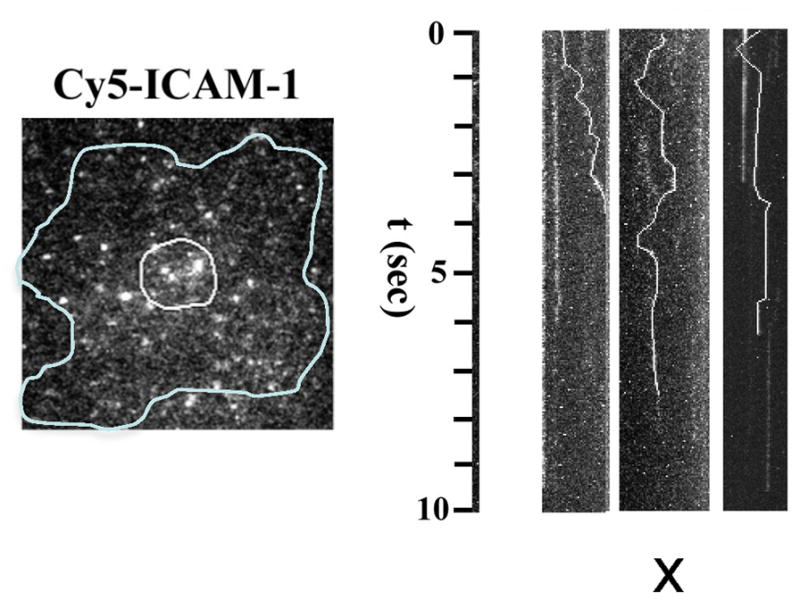
Integrins dynamically equilibrate between three conformational states on cell surfaces. A bent conformation has a closed headpiece. Two extended conformations contain either a closed or an open headpiece. Headpiece opening involves hybrid domain swing-out and a 70 Å separation at the integrin knees, which is conveyed by allostery from the hybrid-proximal end of the βI domain to a 3 Å rearrangement of the ligand-binding site at the opposite end of the βI domain. Both bent-closed and extended-closed integrins have low affinity, whereas extended-open integrin affinity is 10(3) to 10(4) higher. Integrin-mediated adhesion requires the extended-open conformation, which in physiological contexts is stabilized by post-ligand binding events. Integrins thus discriminate between substrate-bound and soluble ligands. Analysis of LFA-1-ICAM-1 interactions in the immunological synapse suggests that bond lifetimes are on the order of seconds, which is consistent with high affinity interactions subjected to cytoskeletal forces that increase the dissociation rate. LFA-1 βI domain antagonists abrogate function in the immunological synapse, further supporting a critical role for high affinity LFA-1.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
References
-
- Luo B-H, Springer TA, Takagi J. High affinity ligand binding by integrins does not involve head separation. J Biol Chem. 2003;278:17185–17189. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
