

The insulator protein CTCF binding sites in the orf73/LANA promoter region of herpesvirus saimiri are involved in conferring episomal stability in latently infected human T cells
- PMID: 22130528
- PMCID: PMC3264369
- DOI: 10.1128/JVI.06295-11
The insulator protein CTCF binding sites in the orf73/LANA promoter region of herpesvirus saimiri are involved in conferring episomal stability in latently infected human T cells
Abstract
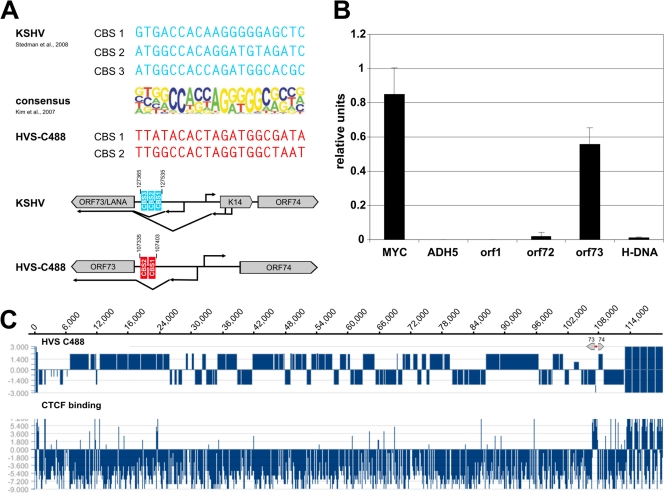
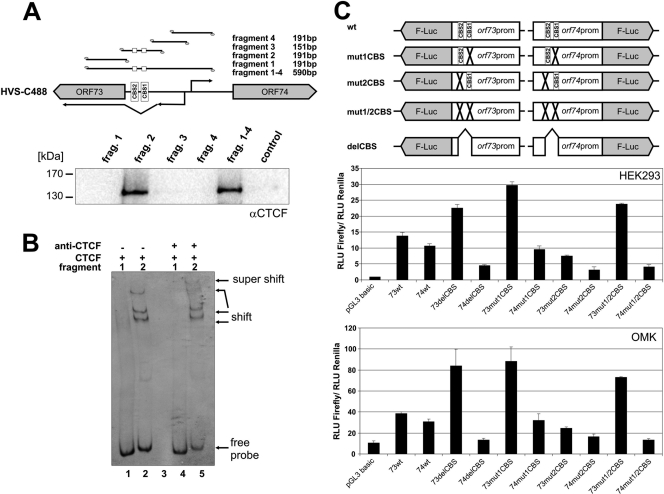
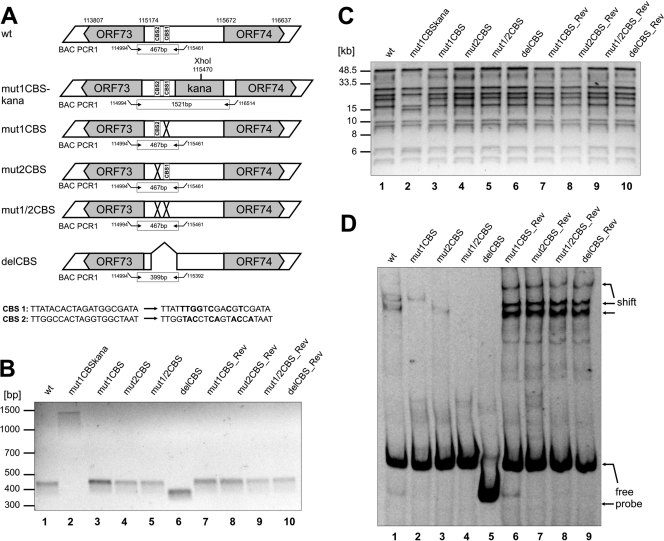
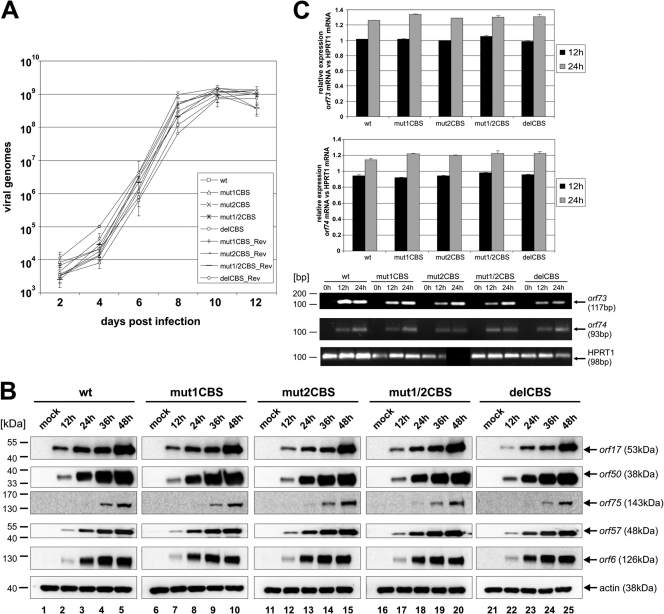
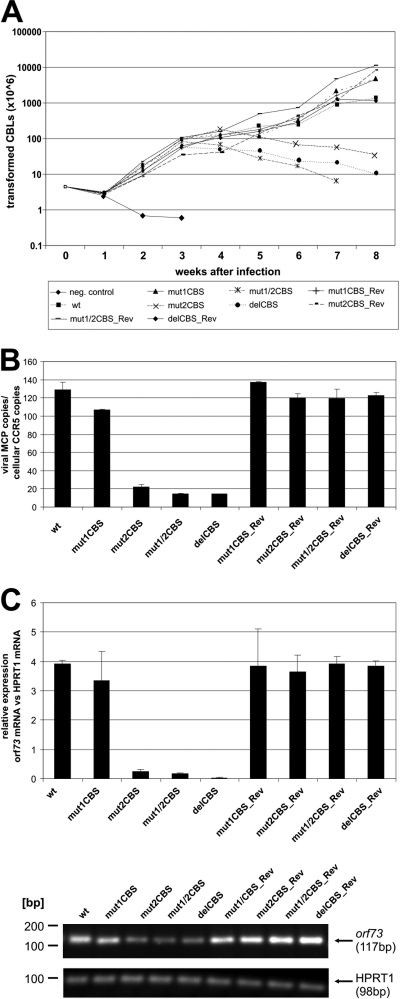
Herpesviruses establish latency in suitable cells of the host organism after a primary lytic infection. Subgroup C strains of herpesvirus saimiri (HVS), a primate gamma-2 herpesvirus, are able to transform human and other primate T lymphocytes to stable growth in vitro. The viral genomes persist as nonintegrated, circular, and histone-associated episomes in the nuclei of those latently infected T cells. Epigenetic modifications of episomes are essential to restrict the transcription during latency to selected viral genes, such as the viral oncogenes stpC/tip and the orf73/LANA. In this study, we describe a genome-wide chromatin immunoprecipitation-on-chip (ChIP-on-chip) analysis to profile the occupancy of CTCF on the latent HVS genome. We then focused on two distinct, conserved CTCF binding sites (CBS) within the orf73/LANA promoter region. Analysis of recombinant viruses harboring deletions or mutations within the CBS indicated that the lytic replication of such viruses is not substantially influenced by CTCF. However, T cells latently infected with CBS mutants were impaired in their proliferation abilities and showed a significantly reduced episomal maintenance. We detected a reduced transcription of the orf73/LANA gene in the T cells, corresponding to the reduced viral genomes; this might contribute to the loss of HVS episomes, as LANA is central in the maintenance of viral episomes in the dividing T cell populations. These data demonstrate that the episomal stability of HVS genomes in latently infected human T cells is dependent on CTCF.
Figures
Similar articles
-
Herpesvirus saimiri episomal persistence is maintained via interaction between open reading frame 73 and the cellular chromosome-associated protein MeCP2.J Virol. 2007 Apr;81(8):4021-32. doi: 10.1128/JVI.02171-06. Epub 2007 Jan 31. J Virol. 2007. PMID: 17267510 Free PMC article.
-
ORF73 of herpesvirus Saimiri strain C488 tethers the viral genome to metaphase chromosomes and binds to cis-acting DNA sequences in the terminal repeats.J Virol. 2003 Dec;77(23):12494-506. doi: 10.1128/jvi.77.23.12494-12506.2003. J Virol. 2003. PMID: 14610173 Free PMC article.
-
Histone modification pattern of the T-cellular Herpesvirus saimiri genome in latency.J Virol. 2007 Mar;81(5):2524-30. doi: 10.1128/JVI.01931-06. Epub 2006 Dec 6. J Virol. 2007. PMID: 17151105 Free PMC article.
-
Herpesvirus saimiri.Philos Trans R Soc Lond B Biol Sci. 2001 Apr 29;356(1408):545-67. doi: 10.1098/rstb.2000.0780. Philos Trans R Soc Lond B Biol Sci. 2001. PMID: 11313011 Free PMC article. Review.
-
Epigenetic regulation of latent Epstein-Barr virus promoters.Biochim Biophys Acta. 2010 Mar-Apr;1799(3-4):228-35. doi: 10.1016/j.bbagrm.2009.10.005. Epub 2009 Oct 22. Biochim Biophys Acta. 2010. PMID: 19853674 Review.
Cited by
-
Efficient generation of human natural killer cell lines by viral transformation.Leukemia. 2014 Jan;28(1):192-5. doi: 10.1038/leu.2013.188. Epub 2013 Jun 21. Leukemia. 2014. PMID: 23787393 No abstract available.
-
Herpesvirus saimiri antagonizes nuclear domain 10-instituted intrinsic immunity via an ORF3-mediated selective degradation of cellular protein Sp100.J Virol. 2012 Apr;86(7):3541-53. doi: 10.1128/JVI.06992-11. Epub 2012 Jan 25. J Virol. 2012. PMID: 22278248 Free PMC article.
-
Targeting CTCF to Control Virus Gene Expression: A Common Theme amongst Diverse DNA Viruses.Viruses. 2015 Jul 6;7(7):3574-85. doi: 10.3390/v7072791. Viruses. 2015. PMID: 26154016 Free PMC article. Review.
-
The IS2 Element Improves Transcription Efficiency of Integration-Deficient Lentiviral Vector Episomes.Mol Ther Nucleic Acids. 2018 Dec 7;13:16-28. doi: 10.1016/j.omtn.2018.08.007. Epub 2018 Aug 18. Mol Ther Nucleic Acids. 2018. PMID: 30227274 Free PMC article.
-
The β-globin Replicator greatly enhances the potential of S/MAR based episomal vectors for gene transfer into human haematopoietic progenitor cells.Sci Rep. 2017 Jan 20;7:40673. doi: 10.1038/srep40673. Sci Rep. 2017. PMID: 28106085 Free PMC article.
References
-
- Calderwood M, White RE, Griffiths RA, Whitehouse A. 2005. Open reading frame 73 is required for herpesvirus saimiri A11-S4 episomal persistence. J. Gen. Virol. 86:2703–2708 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
