

Avidity binding of human adenovirus serotypes 3 and 7 to the membrane cofactor CD46 triggers infection
- PMID: 22130529
- PMCID: PMC3264387
- DOI: 10.1128/JVI.06181-11
Avidity binding of human adenovirus serotypes 3 and 7 to the membrane cofactor CD46 triggers infection
Abstract
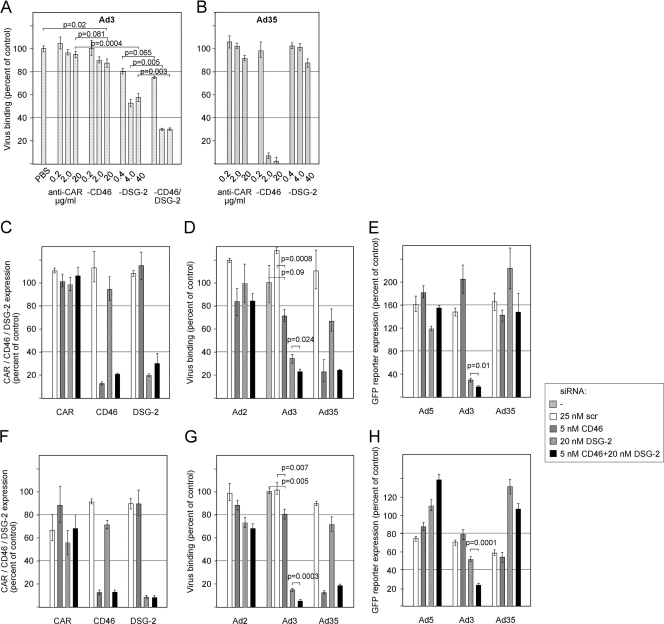
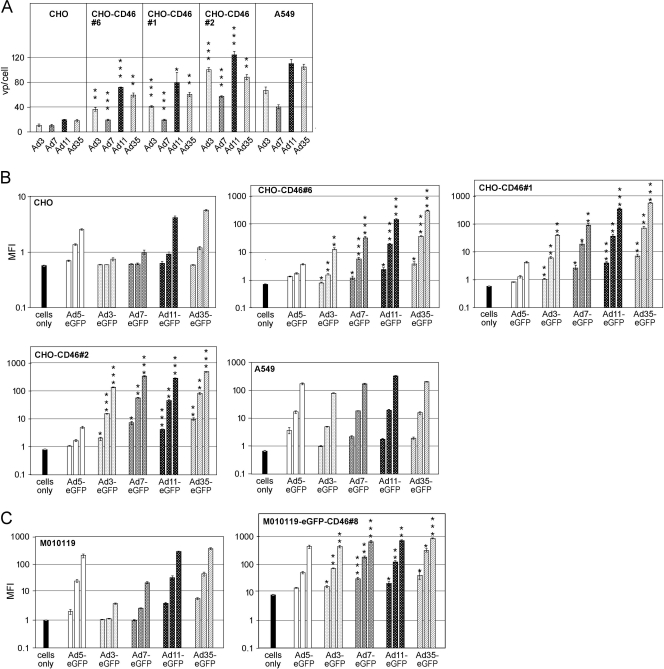
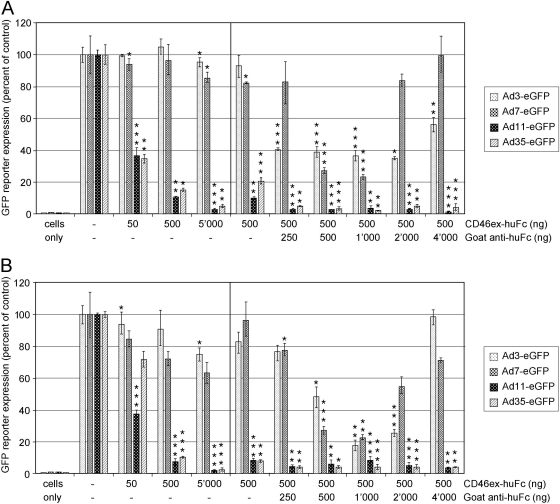
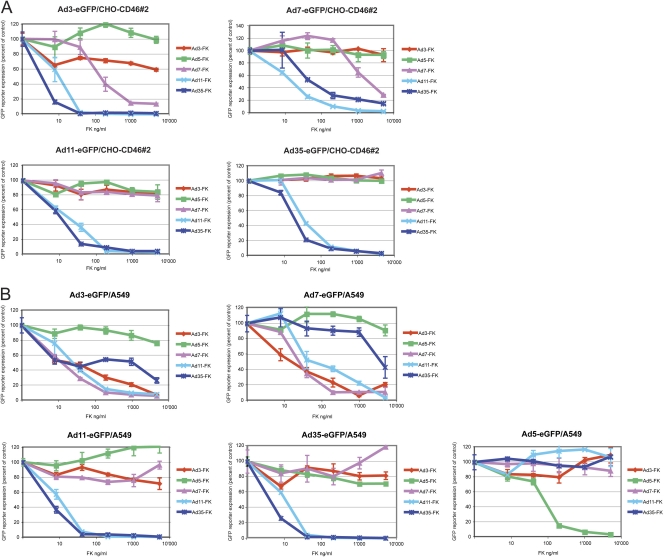
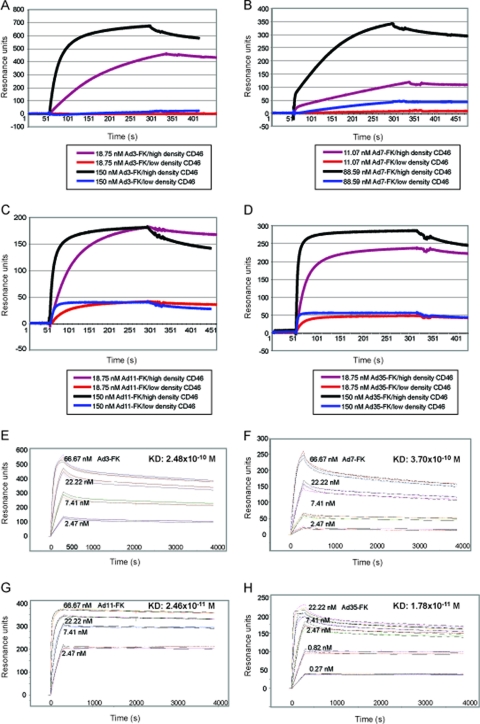
The species B human adenoviruses (HAdVs) infect cells upon attaching to CD46 or desmoglein 2 (DSG-2) by one or several of their 12 fiber knob trimers (FKs). To test whether DSG-2 and CD46 simultaneously serve as virus receptors for adenovirus type 3 (Ad3), we performed individual and combined CD46/DSG-2 loss-of-function studies in human lung A549 and 16HBE14o cells. Our results suggest that in these cells, DSG-2 functions as a major attachment receptor for Ad3, whereas CD46 exerts a minor contribution to virus attachment and uptake in the range of ∼10%. However, in other cells the role of CD46 may be more pronounced depending on, e.g., the expression levels of the receptors. To test if avidity allows Ad3/7 to use CD46 as a receptor, we performed gain-of-function studies. The cell surface levels of ectopically expressed CD46 in CHO or human M010119 melanoma cells lacking DSG-2 positively correlated with Ad3/7 infections, while Ad11/35 infections depended on CD46 but less on CD46 levels. Antibody-cross-linked soluble CD46 blocked Ad3/7/11/35 infections, while soluble CD46 alone blocked Ad11/35 but not Ad3/7. Soluble Ad3/7-FKs poorly inhibited Ad3/7 infection of CHO-CD46 cells, illustrating that Ad3/7-FKs bind with low affinity to CD46. This was confirmed by Biacore studies. Ad3/7-FK binding to immobilized CD46 at low density was not detected, unlike that of Ad11/35-FK. At higher CD46 densities, however, Ad3/7-FK bound to CD46 with only 15-fold-higher dissociation constants than those of Ad11/35-FK. These data show that an avidity mechanism for Ad3/7 binding to CD46 leads to infection of CD46-positive cells.
Figures
Similar articles
-
The human membrane cofactor CD46 is a receptor for species B adenovirus serotype 3.J Virol. 2004 May;78(9):4454-62. doi: 10.1128/jvi.78.9.4454-4462.2004. J Virol. 2004. PMID: 15078926 Free PMC article.
-
Species B adenovirus serotypes 3, 7, 11 and 35 share similar binding sites on the membrane cofactor protein CD46 receptor.J Gen Virol. 2007 Nov;88(Pt 11):2925-2934. doi: 10.1099/vir.0.83142-0. J Gen Virol. 2007. PMID: 17947513
-
Defining the role of CD46, CD80 and CD86 in mediating adenovirus type 3 fiber interactions with host cells.Virology. 2009 Sep 30;392(2):222-9. doi: 10.1016/j.virol.2009.07.010. Epub 2009 Aug 13. Virology. 2009. PMID: 19682720
-
Adenovirus type 11 uses CD46 as a cellular receptor.J Virol. 2003 Sep;77(17):9183-91. doi: 10.1128/jvi.77.17.9183-9191.2003. J Virol. 2003. PMID: 12915534 Free PMC article.
-
Human adenovirus binding to host cell receptors: a structural view.Med Microbiol Immunol. 2020 Jun;209(3):325-333. doi: 10.1007/s00430-019-00645-2. Epub 2019 Nov 29. Med Microbiol Immunol. 2020. PMID: 31784892 Free PMC article. Review.
Cited by
-
Host cell autophagy modulates early stages of adenovirus infections in airway epithelial cells.J Virol. 2013 Feb;87(4):2307-19. doi: 10.1128/JVI.02014-12. Epub 2012 Dec 12. J Virol. 2013. PMID: 23236070 Free PMC article.
-
1. Alternative splicing of viral receptors: A review of the diverse morphologies and physiologies of adenoviral receptors.Recent Res Dev Virol. 2014;9:1-24. Recent Res Dev Virol. 2014. PMID: 25621323 Free PMC article.
-
A Human In Vitro Model to Study Adenoviral Receptors and Virus Cell Interactions.Cells. 2022 Mar 1;11(5):841. doi: 10.3390/cells11050841. Cells. 2022. PMID: 35269463 Free PMC article.
-
Human Desmoglein-2 and Human CD46 Mediate Human Adenovirus Type 55 Infection, but Human Desmoglein-2 Plays the Major Roles.J Virol. 2020 Aug 17;94(17):e00747-20. doi: 10.1128/JVI.00747-20. Print 2020 Aug 17. J Virol. 2020. PMID: 32581096 Free PMC article.
-
Efficacy of CD46-targeting chimeric Ad5/35 adenoviral gene therapy for colorectal cancers.Oncotarget. 2016 Jun 21;7(25):38210-38223. doi: 10.18632/oncotarget.9427. Oncotarget. 2016. PMID: 27203670 Free PMC article.
References
-
- Barton ES, et al. 2001. Junction adhesion molecule is a receptor for reovirus. Cell 104:441–451 - PubMed
-
- Blau DM, Compans RW. 1995. Entry and release of measles virus are polarized in epithelial cells. Virology 210:91–99 - PubMed
-
- Burckhardt CJ, et al. 2011. Drifting motions of the adenovirus receptor CAR and immobile integrins initiate virus uncoating and membrane lytic protein exposure. Cell Host Microbe 10:105–117 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
