

SIRT1 modulates aggregation and toxicity through deacetylation of the androgen receptor in cell models of SBMA
- PMID: 22131404
- PMCID: PMC6088793
- DOI: 10.1523/JNEUROSCI.3958-11.2011
SIRT1 modulates aggregation and toxicity through deacetylation of the androgen receptor in cell models of SBMA
Abstract
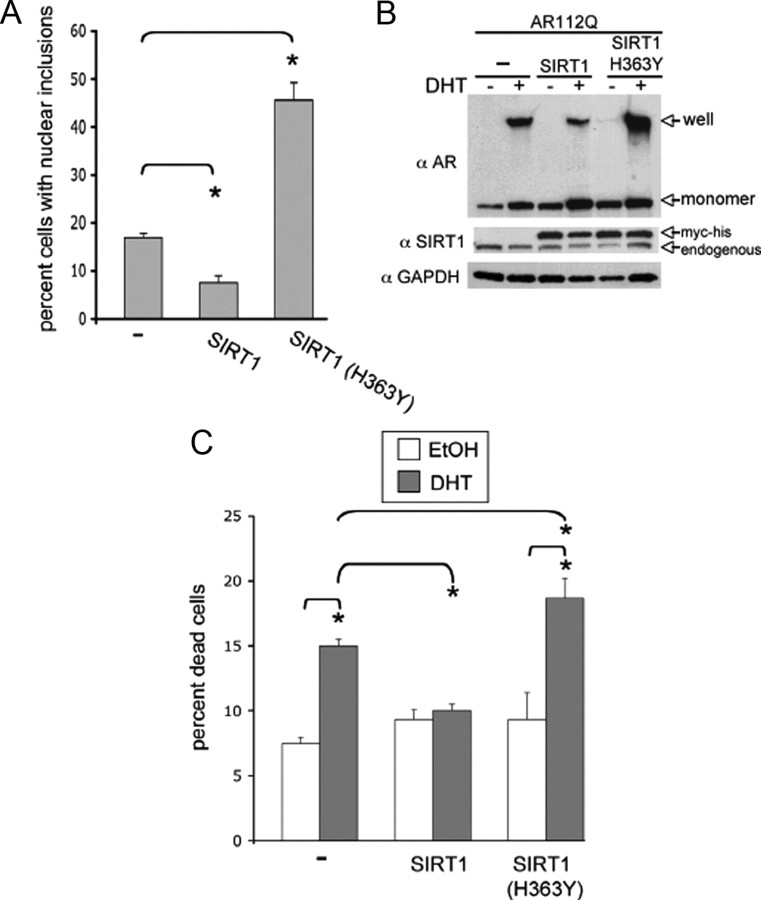
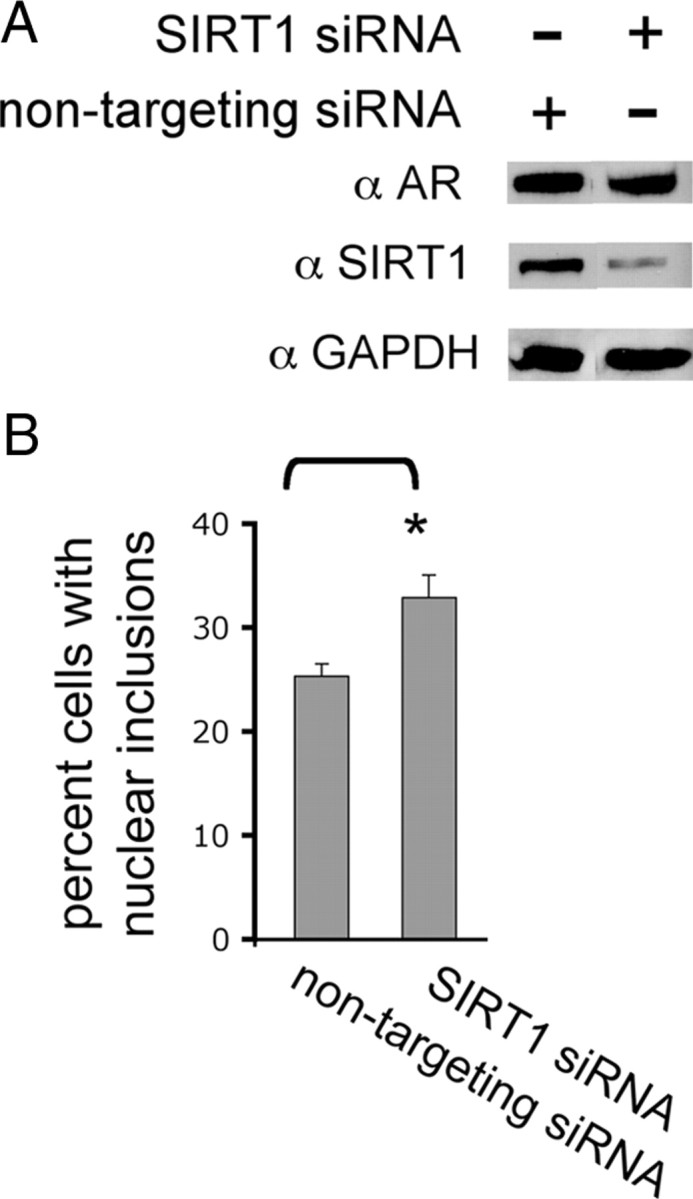
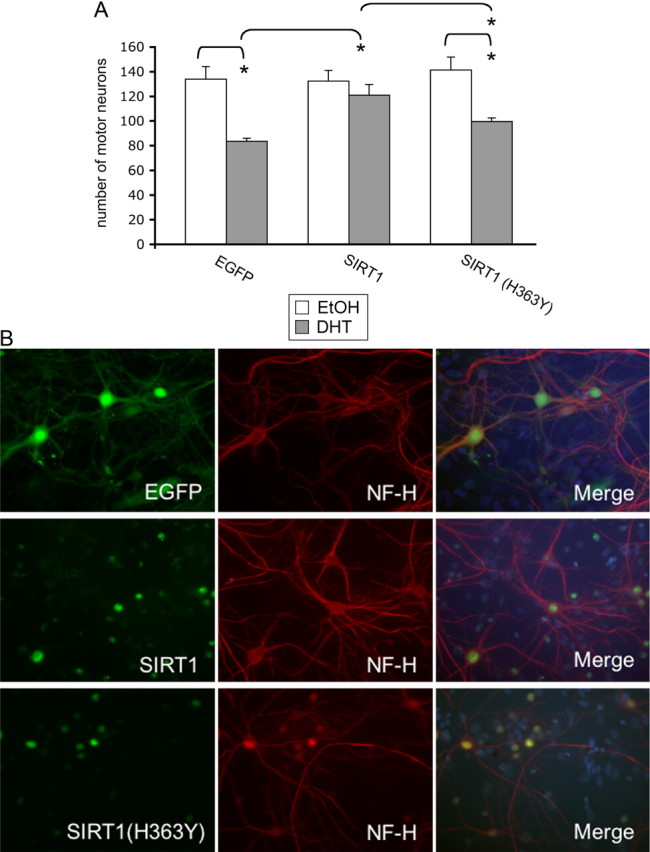
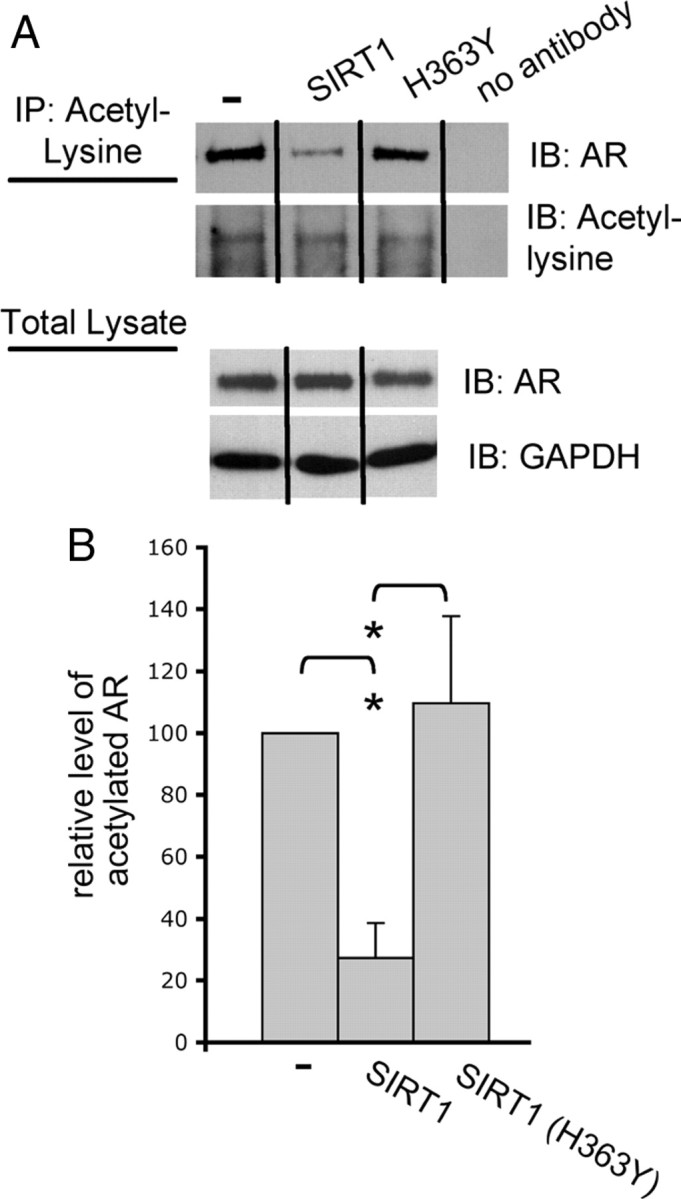
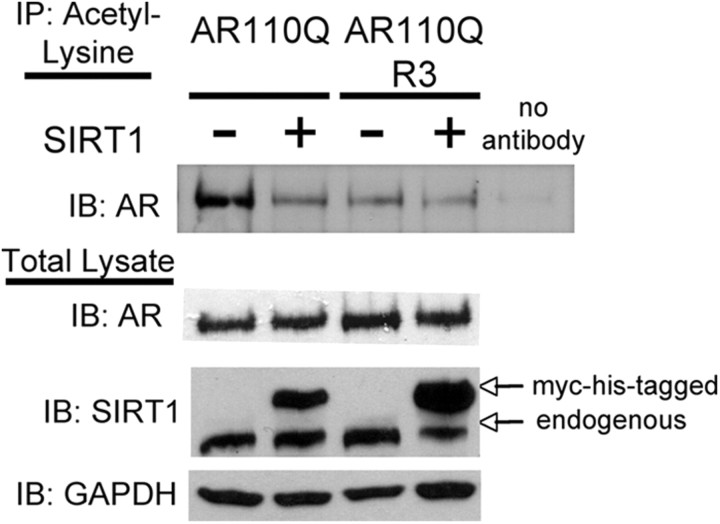
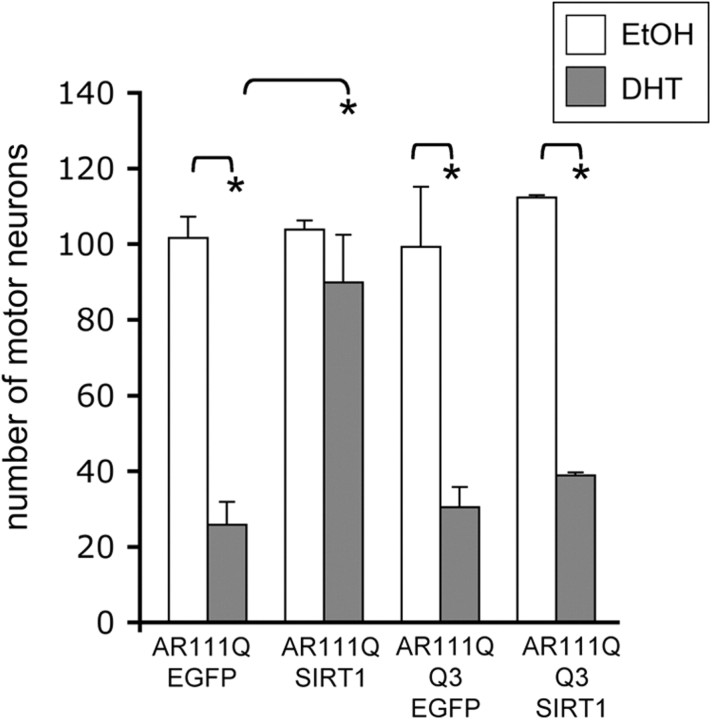
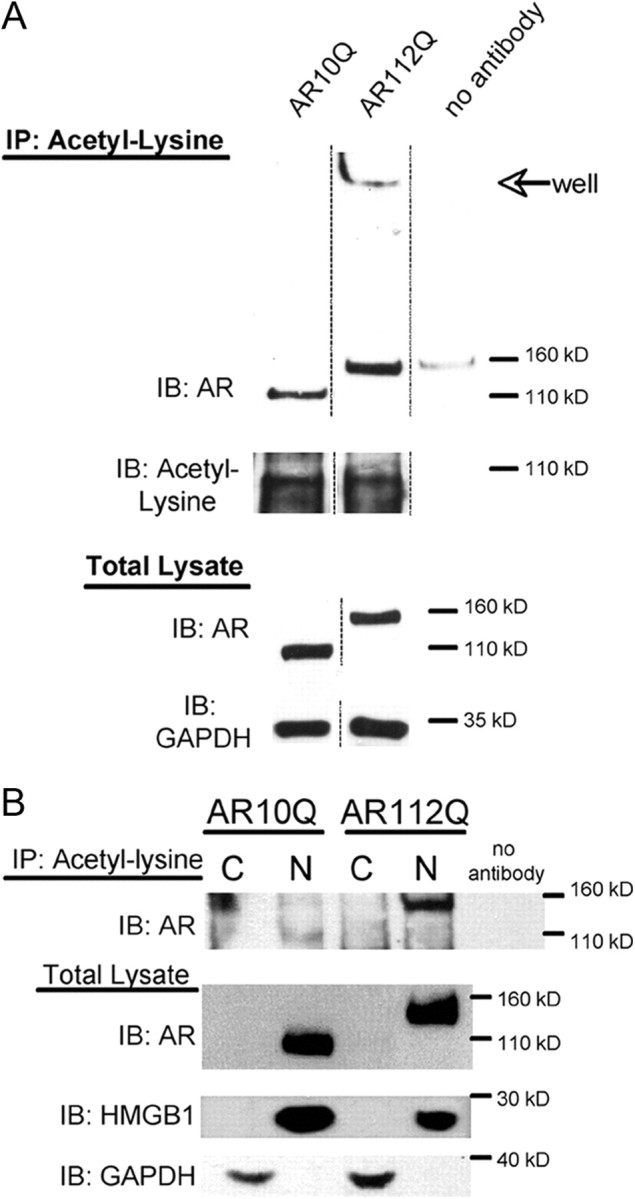
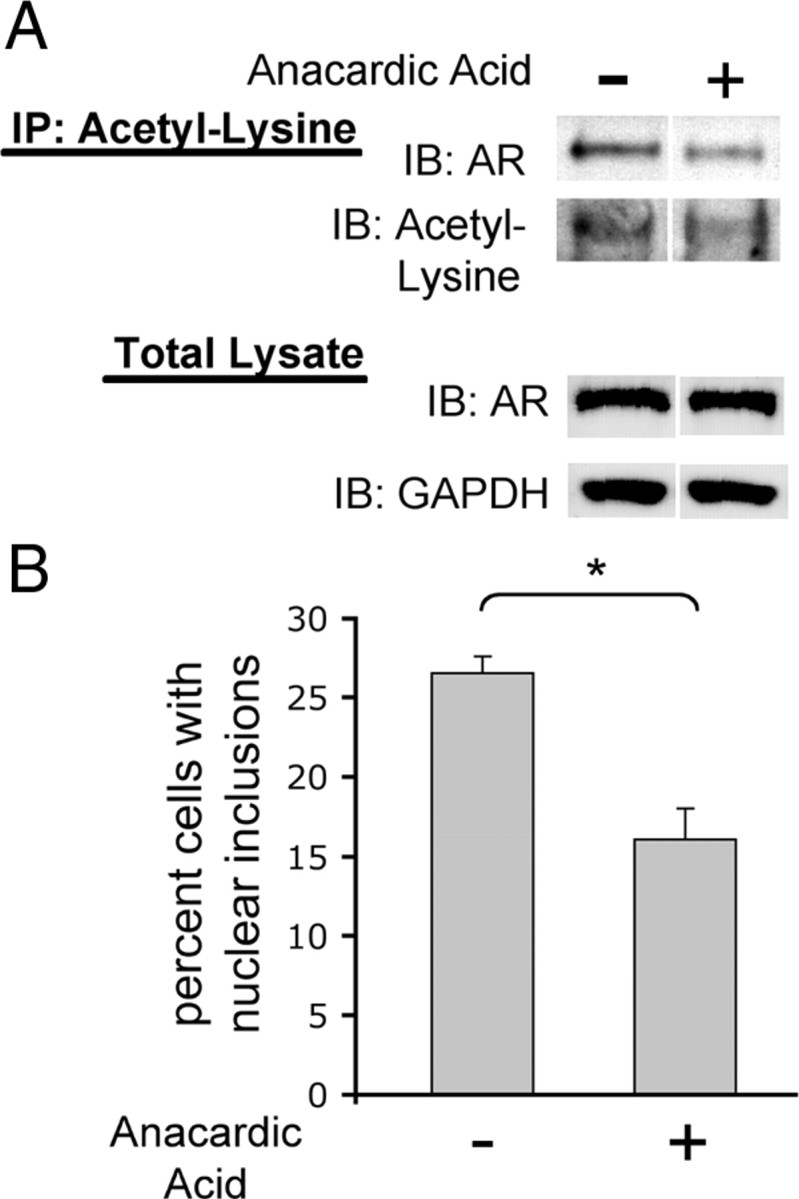
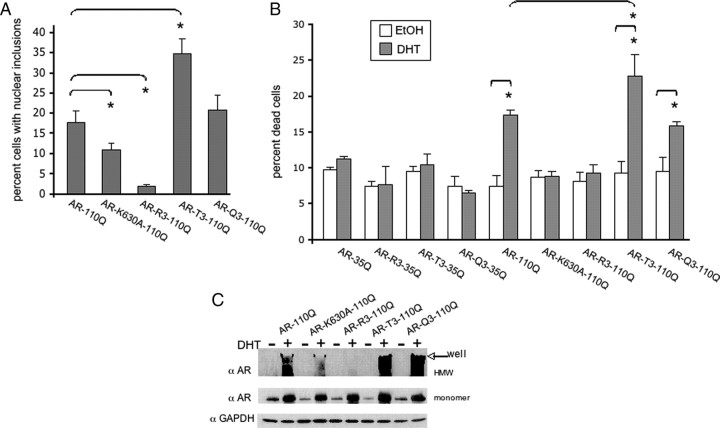
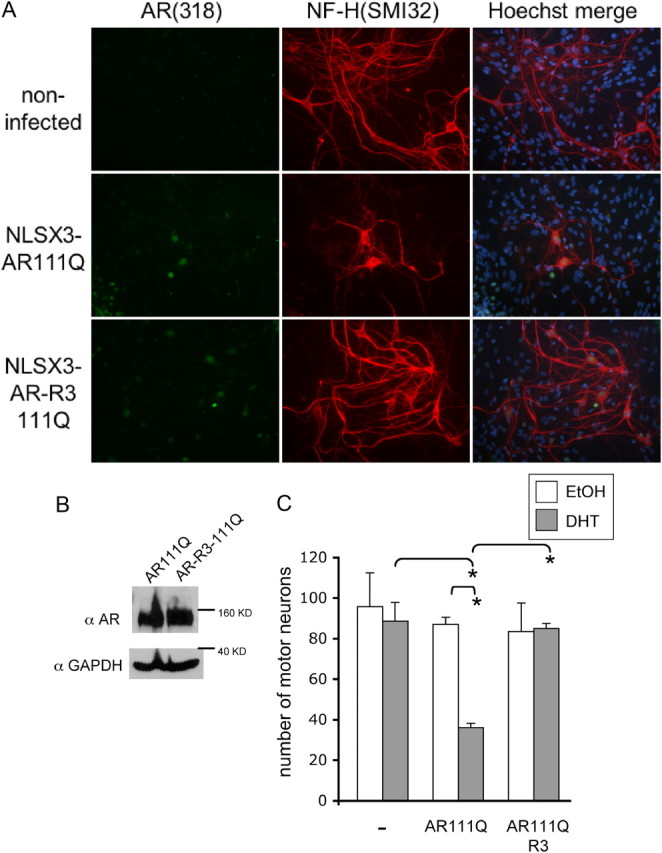
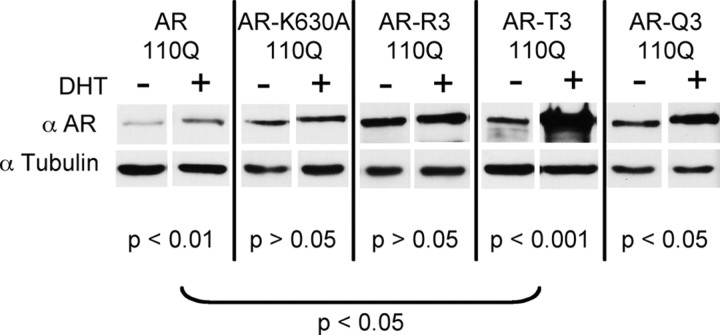
Posttranslational protein modifications can play a major role in disease pathogenesis; phosphorylation, sumoylation, and acetylation modulate the toxicity of a variety of proteotoxic proteins. The androgen receptor (AR) is substantially modified, in response to hormone binding, by phosphorylation, sumoylation, and acetylation; these modifications might thus contribute to DHT-dependent polyglutamine (polyQ)-expanded AR proteotoxicity in spinal and bulbar muscular atrophy (SBMA). SIRT1, a nuclear protein and deacetylase of the AR, is neuroprotective in many neurodegenerative disease models. Our studies reveal that SIRT1 also offers protection against polyQ-expanded AR by deacetylating the AR at lysines 630/632/633. This finding suggested that nuclear AR acetylation plays a role in the aberrant metabolism and toxicity of polyQ-expanded AR. Subsequent studies revealed that the polyQ-expanded AR is hyperacetylated and that pharmacologic reduction of acetylation reduces mutant AR aggregation. Moreover, genetic mutation to inhibit polyQ-expanded AR acetylation of lysines 630/632/633 substantially decreased its aggregation and completely abrogated its toxicity in cell lines and motor neurons. Our studies also reveal one means by which the AR acetylation state likely modifies polyQ-expanded AR metabolism and toxicity, through its effect on DHT-dependent AR stabilization. Overall, our findings reveal a neuroprotective function of SIRT1 that operates through its deacetylation of polyQ-expanded AR and highlight the potential of both SIRT1 and AR acetylation as powerful therapeutic targets in SBMA.
Figures
Similar articles
-
Cytoplasmic retention of polyglutamine-expanded androgen receptor ameliorates disease via autophagy in a mouse model of spinal and bulbar muscular atrophy.Hum Mol Genet. 2009 Jun 1;18(11):1937-50. doi: 10.1093/hmg/ddp115. Epub 2009 Mar 11. Hum Mol Genet. 2009. PMID: 19279159 Free PMC article.
-
Mutant androgen receptor induces neurite loss and senescence independently of ARE binding in a neuronal model of SBMA.Proc Natl Acad Sci U S A. 2024 Jul 16;121(29):e2321408121. doi: 10.1073/pnas.2321408121. Epub 2024 Jul 8. Proc Natl Acad Sci U S A. 2024. PMID: 38976730 Free PMC article.
-
Loss of endogenous androgen receptor protein accelerates motor neuron degeneration and accentuates androgen insensitivity in a mouse model of X-linked spinal and bulbar muscular atrophy.Hum Mol Genet. 2006 Jul 15;15(14):2225-38. doi: 10.1093/hmg/ddl148. Epub 2006 Jun 13. Hum Mol Genet. 2006. PMID: 16772330
-
Spinal and bulbar muscular atrophy: ligand-dependent pathogenesis and therapeutic perspectives.J Mol Med (Berl). 2004 May;82(5):298-307. doi: 10.1007/s00109-004-0530-7. Epub 2004 Feb 27. J Mol Med (Berl). 2004. PMID: 15133611 Review.
-
Transgenic mouse models of spinal and bulbar muscular atrophy (SBMA).Cytogenet Genome Res. 2003;100(1-4):243-51. doi: 10.1159/000072860. Cytogenet Genome Res. 2003. PMID: 14526186 Review.
Cited by
-
SIRT1 Promotes Neuronal Fortification in Neurodegenerative Diseases through Attenuation of Pathological Hallmarks and Enhancement of Cellular Lifespan.Curr Neuropharmacol. 2021;19(7):1019-1037. doi: 10.2174/1570159X18666200729111744. Curr Neuropharmacol. 2021. PMID: 32727328 Free PMC article. Review.
-
A survey of protein interactions and posttranslational modifications that influence the polyglutamine diseases.Front Mol Neurosci. 2022 Sep 14;15:974167. doi: 10.3389/fnmol.2022.974167. eCollection 2022. Front Mol Neurosci. 2022. PMID: 36187346 Free PMC article. Review.
-
Polyglutamine androgen receptor-mediated neuromuscular disease.Cell Mol Life Sci. 2016 Nov;73(21):3991-9. doi: 10.1007/s00018-016-2275-1. Epub 2016 May 17. Cell Mol Life Sci. 2016. PMID: 27188284 Free PMC article. Review.
-
Exploring the Role of Posttranslational Modifications in Spinal and Bulbar Muscular Atrophy.Front Mol Neurosci. 2022 Jun 3;15:931301. doi: 10.3389/fnmol.2022.931301. eCollection 2022. Front Mol Neurosci. 2022. PMID: 35726299 Free PMC article. Review.
-
p62/SQSTM1 differentially removes the toxic mutant androgen receptor via autophagy and inclusion formation in a spinal and bulbar muscular atrophy mouse model.J Neurosci. 2013 May 1;33(18):7710-27. doi: 10.1523/JNEUROSCI.3021-12.2013. J Neurosci. 2013. PMID: 23637164 Free PMC article.
References
-
- Araki T, Sasaki Y, Milbrandt J. Increased nuclear NAD biosynthesis and SIRT1 activation prevent axonal degeneration. Science. 2004;305:1010–1013. - PubMed
-
- Bouras T, Fu M, Sauve AA, Wang F, Quong AA, Perkins ND, Hay RT, Gu W, Pestell RG. SIRT1 deacetylation and repression of p300 involves lysine residues 1020/1024 within the cell cycle regulatory domain 1. J Biol Chem. 2005;280:10264–10276. - PubMed
-
- Fu M, Wang C, Reutens AT, Wang J, Angeletti RH, Siconolfi-Baez L, Ogryzko V, Avantaggiati ML, Pestell RG. p300 and p300/cAMP-response element-binding protein-associated factor acetylate the androgen receptor at sites governing hormone-dependent transactivation. J Biol Chem. 2000;275:20853–20860. - PubMed
-
- Fu M, Wang C, Wang J, Zhang X, Sakamaki T, Yeung YG, Chang C, Hopp T, Fuqua SA, Jaffray E, Hay RT, Palvimo JJ, Jänne OA, Pestell RG. Androgen receptor acetylation governs trans activation and MEKK1-induced apoptosis without affecting in vitro sumoylation and trans-repression function. Mol Cell Biol. 2002;22:3373–3388. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials