

Structural analysis of prolyl oligopeptidases using molecular docking and dynamics: insights into conformational changes and ligand binding
- PMID: 22132071
- PMCID: PMC3223163
- DOI: 10.1371/journal.pone.0026251
Structural analysis of prolyl oligopeptidases using molecular docking and dynamics: insights into conformational changes and ligand binding
Abstract
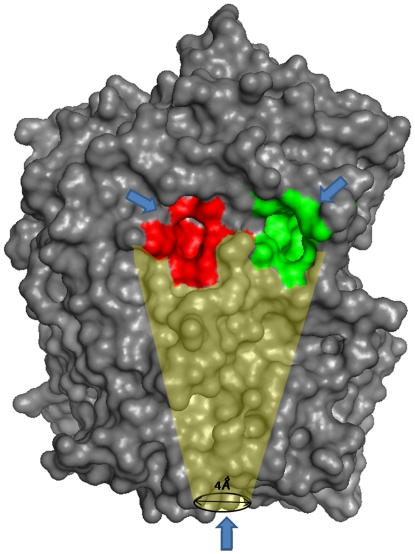
Prolyl oligopeptidase (POP) is considered as an important pharmaceutical target for the treatment of numerous diseases. Despite enormous studies on various aspects of POPs structure and function still some of the questions are intriguing like conformational dynamics of the protein and interplay between ligand entry/egress. Here, we have used molecular modeling and docking based approaches to unravel questions like differences in ligand binding affinities in three POP species (porcine, human and A. thaliana). Despite high sequence and structural similarity, they possess different affinities for the ligands. Interestingly, human POP was found to be more specific, selective and incapable of binding to a few planar ligands which showed extrapolation of porcine POP in human context is more complicated. Possible routes for substrate entry and product egress were also investigated by detailed analyses of molecular dynamics (MD) simulations for the three proteins. Trajectory analysis of bound and unbound forms of three species showed differences in conformational dynamics, especially variations in β-propeller pore size, which was found to be hidden by five lysine residues present on blades one and seven. During simulation, β-propeller pore size was increased by ∼2 Å in porcine ligand-bound form which might act as a passage for smaller product movement as free energy barrier was reduced, while there were no significant changes in human and A. thaliana POPs. We also suggest that these differences in pore size could lead to fundamental differences in mode of product egress among three species. This analysis also showed some functionally important residues which can be used further for in vitro mutagenesis and inhibitor design. This study can help us in better understanding of the etiology of POPs in several neurodegenerative diseases.
Conflict of interest statement
Figures










Similar articles
-
Decoding the structural events in substrate-gating mechanism of eukaryotic prolyl oligopeptidase using normal mode analysis and molecular dynamics simulations.Proteins. 2014 Jul;82(7):1428-43. doi: 10.1002/prot.24511. Epub 2014 Feb 6. Proteins. 2014. PMID: 24500901
-
The structure and molecular dynamics of prolyl oligopeptidase from Microbulbifer arenaceous provide insights into catalytic and regulatory mechanisms.Acta Crystallogr D Struct Biol. 2022 Jun 1;78(Pt 6):735-751. doi: 10.1107/S2059798322004247. Epub 2022 May 9. Acta Crystallogr D Struct Biol. 2022. PMID: 35647921
-
Substrate-like novel inhibitors of prolyl specific oligo peptidase for neurodegenerative disorders.J Biomol Struct Dyn. 2024 Oct;42(16):8454-8472. doi: 10.1080/07391102.2023.2246577. Epub 2023 Aug 22. J Biomol Struct Dyn. 2024. PMID: 37608559
-
Structure, function and biological relevance of prolyl oligopeptidase.Curr Protein Pept Sci. 2008 Feb;9(1):96-107. doi: 10.2174/138920308783565723. Curr Protein Pept Sci. 2008. PMID: 18336325 Review.
-
Prolyl endopeptidases.Cell Mol Life Sci. 2007 Feb;64(3):345-55. doi: 10.1007/s00018-006-6317-y. Cell Mol Life Sci. 2007. PMID: 17160352 Free PMC article. Review.
Cited by
-
Reciprocal influence of protein domains in the cold-adapted acyl aminoacyl peptidase from Sporosarcina psychrophila.PLoS One. 2013;8(2):e56254. doi: 10.1371/journal.pone.0056254. Epub 2013 Feb 15. PLoS One. 2013. PMID: 23457536 Free PMC article.
-
Mechanism of Action of Prolyl Oligopeptidase (PREP) in Degenerative Brain Diseases: Has Peptidase Activity Only a Modulatory Role on the Interactions of PREP with Proteins?Front Aging Neurosci. 2017 Feb 14;9:27. doi: 10.3389/fnagi.2017.00027. eCollection 2017. Front Aging Neurosci. 2017. PMID: 28261087 Free PMC article.
-
Oroxylin A inhibits hemolysis via hindering the self-assembly of α-hemolysin heptameric transmembrane pore.PLoS Comput Biol. 2013;9(1):e1002869. doi: 10.1371/journal.pcbi.1002869. Epub 2013 Jan 17. PLoS Comput Biol. 2013. PMID: 23349625 Free PMC article.
-
Crystal Structure and Conformational Dynamics of Pyrococcus furiosus Prolyl Oligopeptidase.Biochemistry. 2019 Mar 26;58(12):1616-1626. doi: 10.1021/acs.biochem.9b00031. Epub 2019 Mar 5. Biochemistry. 2019. PMID: 30786206 Free PMC article.
-
Antibody Cocktail Exhibits Broad Neutralization Activity Against SARS-CoV-2 and SARS-CoV-2 Variants.Virol Sin. 2021 Oct;36(5):934-947. doi: 10.1007/s12250-021-00409-4. Epub 2021 Jul 5. Virol Sin. 2021. PMID: 34224110 Free PMC article.
References
-
- Serrano SMT, Maroun RC. Snake venom serine proteinases: sequence homology vs. substrate specificity, a paradox to be solved. Toxicon. 2005;45(8):1115–1132. - PubMed
-
- Walter R, Shlank H, Glass JD, Schwartz IL, Kerenyi TD. Leucylglycinamide released from oxytocin by human uterine enzyme. Science. 1971;173(999):827–829. - PubMed
-
- Walter R, Yoshimoto T. Postproline cleaving enzyme: kinetic studies of size and stereospecificity of its active site. Biochemistry. 1978;17(20):4139–4144. - PubMed
-
- Maes M, Goossens F, Scharpé S, Meltzer HY, D'Hondt P, et al. Lower serum prolyl endopeptidase enzyme activity in major depression: further evidence that peptidases play a role in the pathophysiology of depression. Biol Psychiatry. 1994;35(8):545–552. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
