

Contribution of bacterial outer membrane vesicles to innate bacterial defense
- PMID: 22133164
- PMCID: PMC3248377
- DOI: 10.1186/1471-2180-11-258
Contribution of bacterial outer membrane vesicles to innate bacterial defense
Abstract
Background: Outer membrane vesicles (OMVs) are constitutively produced by Gram-negative bacteria throughout growth and have proposed roles in virulence, inflammation, and the response to envelope stress. Here we investigate outer membrane vesiculation as a bacterial mechanism for immediate short-term protection against outer membrane acting stressors. Antimicrobial peptides as well as bacteriophage were used to examine the effectiveness of OMV protection.
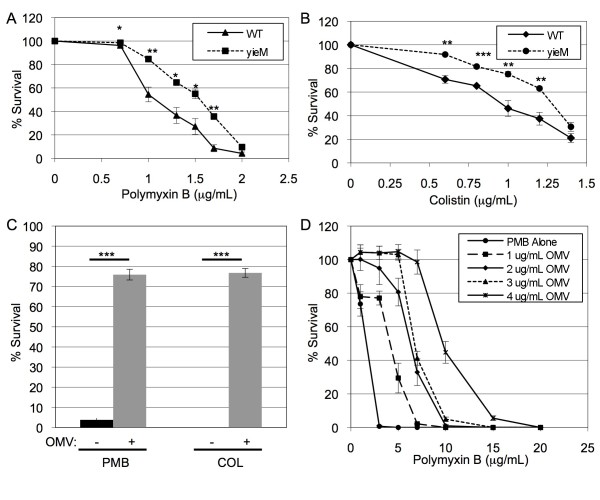
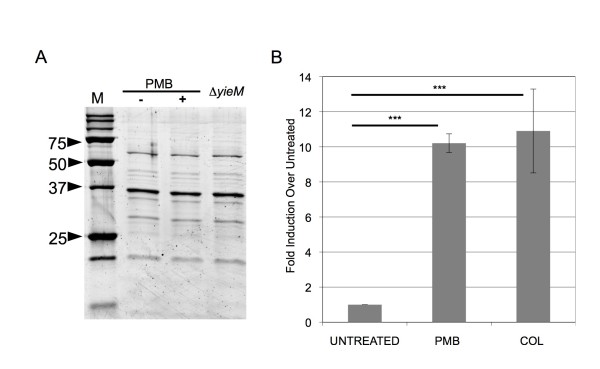
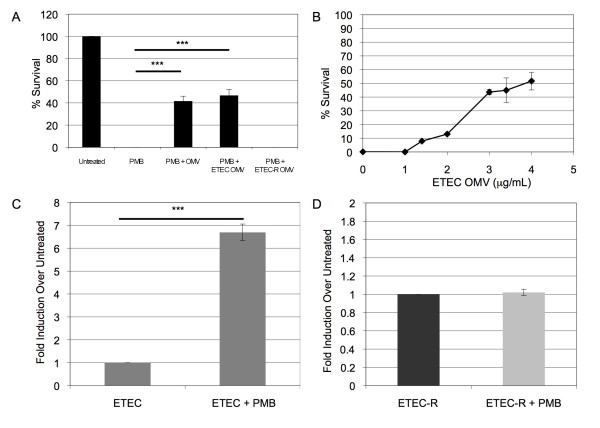
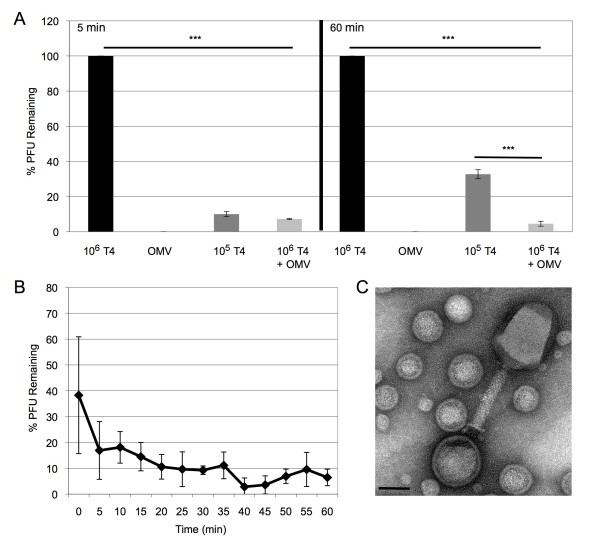
Results: We found that a hyper-vesiculating mutant of Escherichia coli survived treatment by antimicrobial peptides (AMPs) polymyxin B and colistin better than the wild-type. Supplementation of E. coli cultures with purified outer membrane vesicles provided substantial protection against AMPs, and AMPs significantly induced vesiculation. Vesicle-mediated protection and induction of vesiculation were also observed for a human pathogen, enterotoxigenic E. coli (ETEC), challenged with polymyxin B. When ETEC with was incubated with low concentrations of vesicles concomitant with polymyxin B treatment, bacterial survival increased immediately, and the culture gained resistance to polymyxin B. By contrast, high levels of vesicles also provided immediate protection but prevented acquisition of resistance. Co-incubation of T4 bacteriophage and OMVs showed fast, irreversible binding. The efficiency of T4 infection was significantly reduced by the formation of complexes with the OMVs.
Conclusions: These data reveal a role for OMVs in contributing to innate bacterial defense by adsorption of antimicrobial peptides and bacteriophage. Given the increase in vesiculation in response to the antimicrobial peptides, and loss in efficiency of infection with the T4-OMV complex, we conclude that OMV production may be an important factor in neutralizing environmental agents that target the outer membrane of Gram-negative bacteria.
© 2011 Manning and Kuehn; licensee BioMed Central Ltd.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
