

Hyperactivation of oxidative mitochondrial metabolism in epithelial cancer cells in situ: visualizing the therapeutic effects of metformin in tumor tissue
- PMID: 22134189
- PMCID: PMC3272287
- DOI: 10.4161/cc.10.23.18151
Hyperactivation of oxidative mitochondrial metabolism in epithelial cancer cells in situ: visualizing the therapeutic effects of metformin in tumor tissue
Abstract
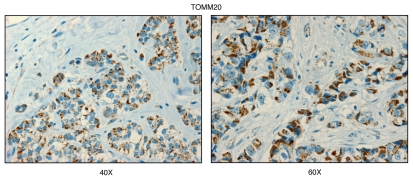
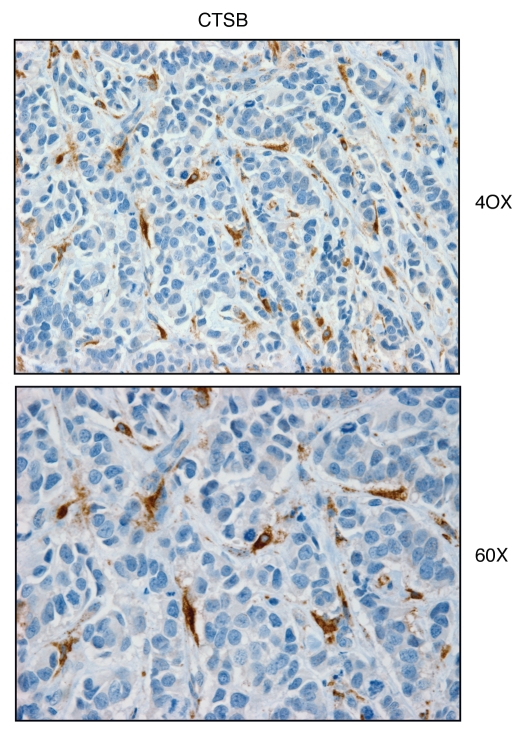
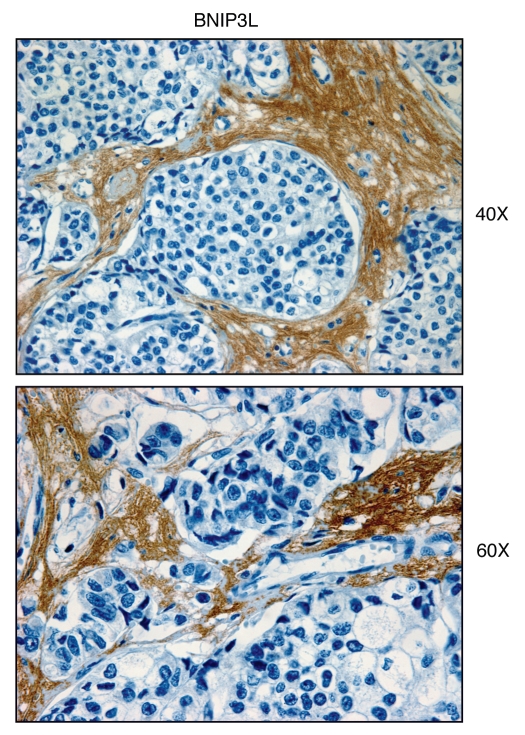
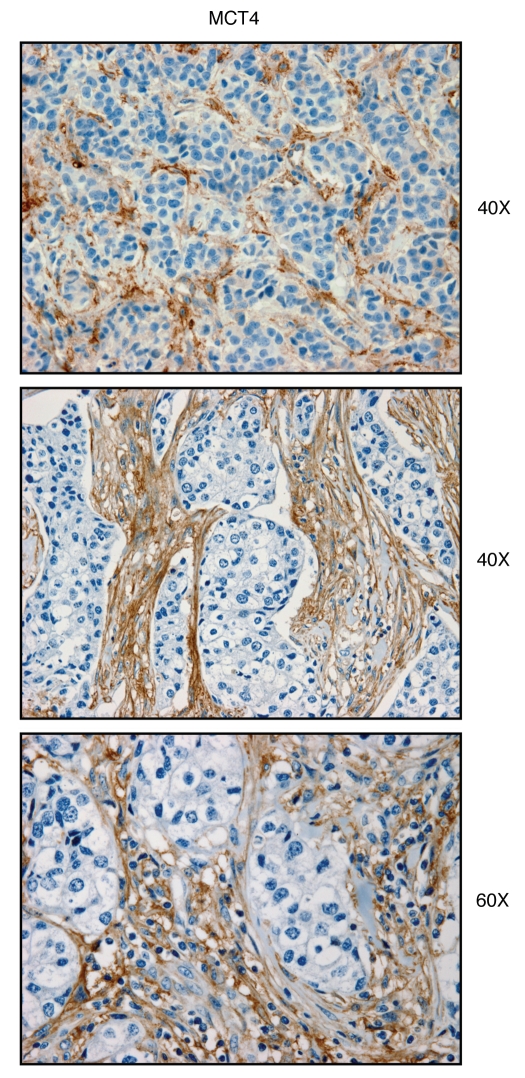
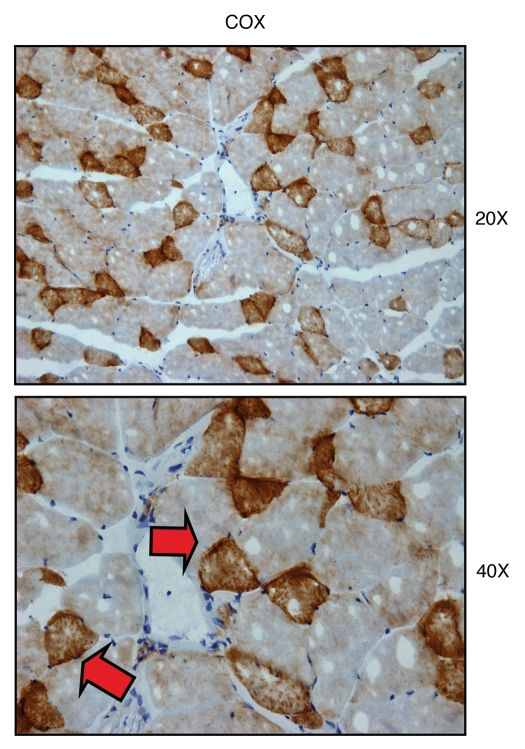
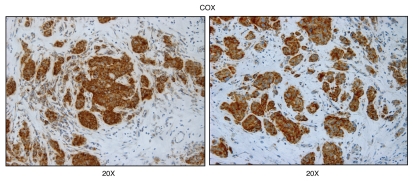
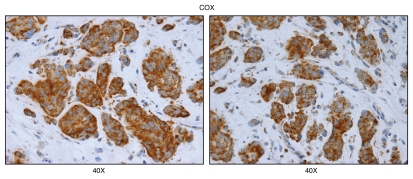
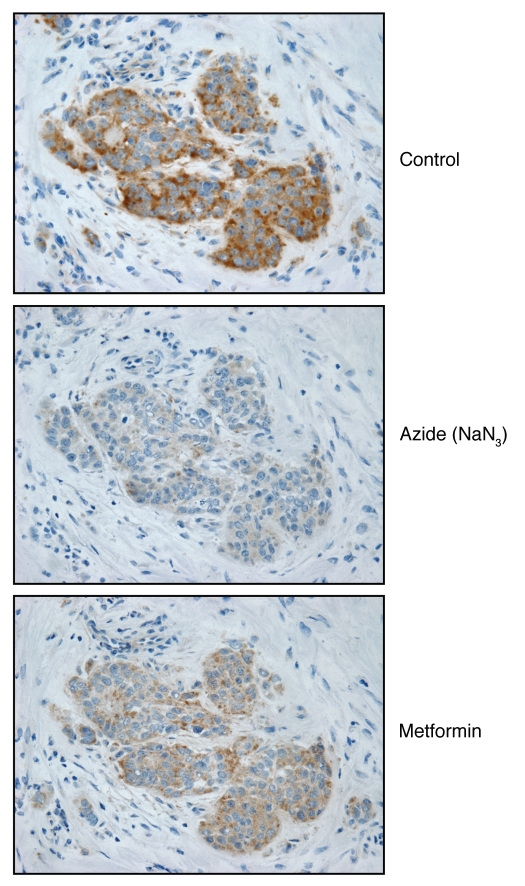
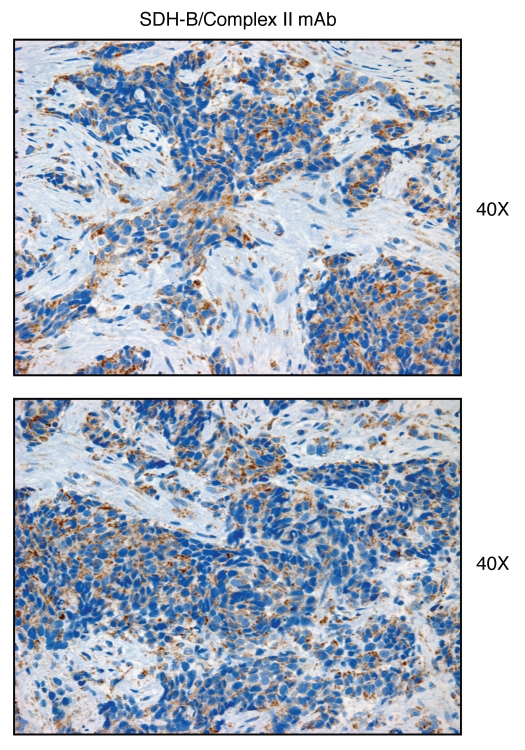
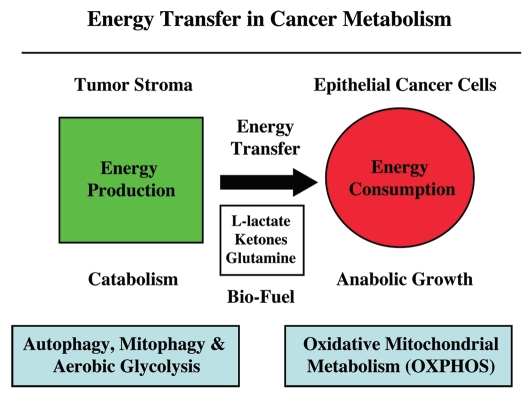
We have recently proposed a new mechanism for explaining energy transfer in cancer metabolism. In this scenario, cancer cells behave as metabolic parasites, by extracting nutrients from normal host cells, such as fibroblasts, via the secretion of hydrogen peroxide as the initial trigger. Oxidative stress in the tumor microenvironment then leads to autophagy-driven catabolism, mitochondrial dys-function, and aerobic glycolysis. This, in turn, produces high-energy nutrients (such as L-lactate, ketones, and glutamine) that drive the anabolic growth of tumor cells, via oxidative mitochondrial metabolism. A logical prediction of this new "parasitic" cancer model is that tumor-associated fibroblasts should show evidence of mitochondrial dys-function (mitophagy and aerobic glycolysis). In contrast, epithelial cancer cells should increase their oxidative mitochondrial capacity. To further test this hypothesis, here we subjected frozen sections from human breast tumors to a staining procedure that only detects functional mitochondria. This method detects the in situ enzymatic activity of cytochrome C oxidase (COX), also known as Complex IV. Remarkably, cancer cells show an over-abundance of COX activity, while adjacent stromal cells remain essentially negative. Adjacent normal ductal epithelial cells also show little or no COX activity, relative to epithelial cancer cells. Thus, oxidative mitochondrial activity is selectively amplified in cancer cells. Although COX activity staining has never been applied to cancer tissues, it could now be used routinely to distinguish cancer cells from normal cells, and to establish negative margins during cancer surgery. Similar results were obtained with NADH activity staining, which measures Complex I activity, and succinate dehydrogenase (SDH) activity staining, which measures Complex II activity. COX and NADH activities were blocked by electron transport inhibitors, such as Metformin. This has mechanistic and clinical implications for using Metformin as an anti-cancer drug, both for cancer therapy and chemo-prevention. We also immuno-stained human breast cancers for a series of well-established protein biomarkers of metabolism. More specifically, we now show that cancer-associated fibroblasts over-express markers of autophagy (cathepsin B), mitophagy (BNIP3L), and aerobic glycolysis (MCT4). Conversely, epithelial cancer cells show the over-expression of a mitochondrial membrane marker (TOMM20), as well as key components of Complex IV (MT-CO1) and Complex II (SDH-B). We also validated our observations using a bioinformatics approach with data from > 2,000 breast cancer patients, which showed the transcriptional upregulation of mitochondrial oxidative phosphorylation (OXPHOS) in human breast tumors (p < 10(-20)), and a specific association with metastasis. Therefore, upregulation of OXPHOS in epithelial tumor cells is a common feature of human breast cancers. In summary, our data provide the first functional in vivo evidence that epithelial cancer cells perform enhanced mitochondrial oxidative phosphorylation, allowing them to produce high amounts of ATP. Thus, we believe that mitochondria are both the "powerhouse" and "Achilles' heel" of cancer cells.
Figures




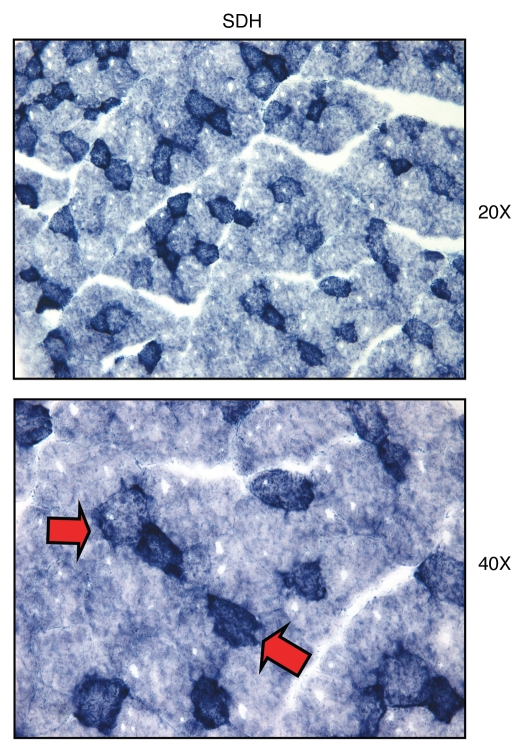
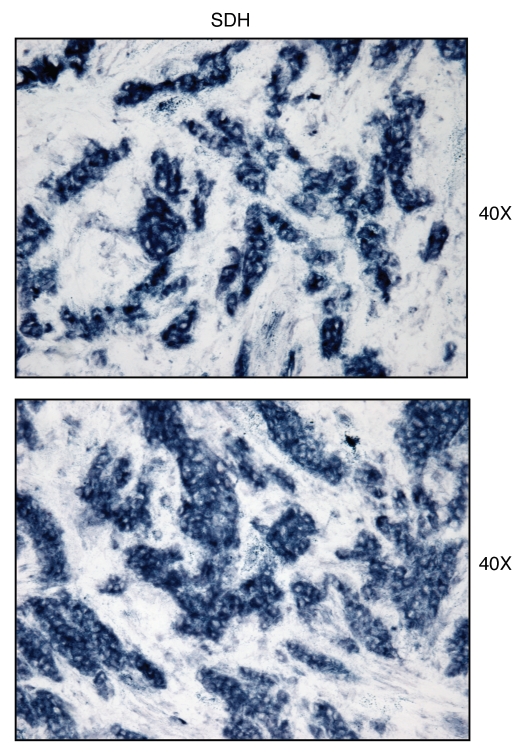
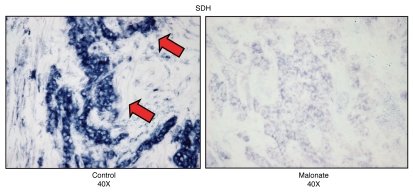
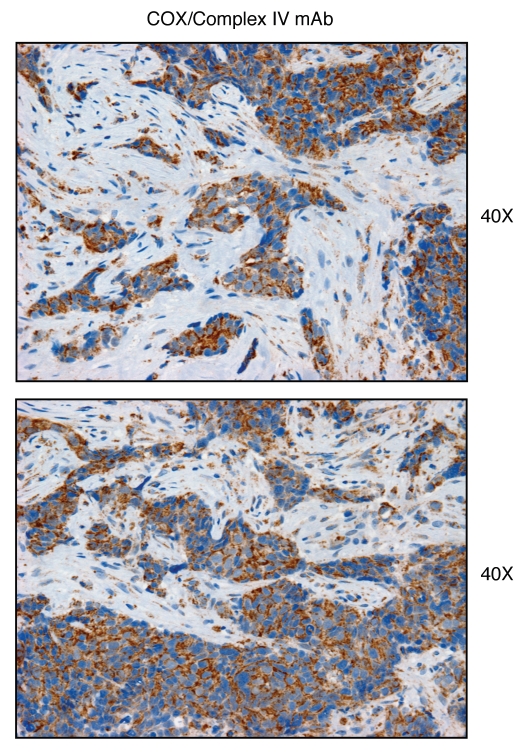

Comment in
-
Metabolic compartments in tumor tissue: implications for therapy.Cell Cycle. 2012 Jan 1;11(1):13-4. doi: 10.4161/cc.11.1.18729. Epub 2012 Jan 1. Cell Cycle. 2012. PMID: 22193036 No abstract available.
Similar articles
-
Cancer metabolism, stemness and tumor recurrence: MCT1 and MCT4 are functional biomarkers of metabolic symbiosis in head and neck cancer.Cell Cycle. 2013 May 1;12(9):1371-84. doi: 10.4161/cc.24092. Epub 2013 Apr 10. Cell Cycle. 2013. PMID: 23574725 Free PMC article.
-
Energy transfer in "parasitic" cancer metabolism: mitochondria are the powerhouse and Achilles' heel of tumor cells.Cell Cycle. 2011 Dec 15;10(24):4208-16. doi: 10.4161/cc.10.24.18487. Epub 2011 Dec 15. Cell Cycle. 2011. PMID: 22033146 Free PMC article.
-
Mitochondrial metabolism in cancer metastasis: visualizing tumor cell mitochondria and the "reverse Warburg effect" in positive lymph node tissue.Cell Cycle. 2012 Apr 1;11(7):1445-54. doi: 10.4161/cc.19841. Epub 2012 Apr 1. Cell Cycle. 2012. PMID: 22395432 Free PMC article.
-
Tumor microenvironment and metabolic synergy in breast cancers: critical importance of mitochondrial fuels and function.Semin Oncol. 2014 Apr;41(2):195-216. doi: 10.1053/j.seminoncol.2014.03.002. Epub 2014 Mar 5. Semin Oncol. 2014. PMID: 24787293 Review.
-
Understanding the Warburg effect and the prognostic value of stromal caveolin-1 as a marker of a lethal tumor microenvironment.Breast Cancer Res. 2011 Jul 8;13(4):213. doi: 10.1186/bcr2892. Breast Cancer Res. 2011. PMID: 21867571 Free PMC article. Review.
Cited by
-
The C30-Modulation of Betulinic Acid Using 1,2,4-Triazole: A Promising Strategy for Increasing Its Antimelanoma Cytotoxic Potential.Molecules. 2022 Nov 12;27(22):7807. doi: 10.3390/molecules27227807. Molecules. 2022. PMID: 36431906 Free PMC article.
-
Targeting the Metabolic Paradigms in Cancer and Diabetes.Biomedicines. 2024 Jan 17;12(1):211. doi: 10.3390/biomedicines12010211. Biomedicines. 2024. PMID: 38255314 Free PMC article. Review.
-
Redox biology and the interface between bioenergetics, autophagy and circadian control of metabolism.Free Radic Biol Med. 2016 Nov;100:94-107. doi: 10.1016/j.freeradbiomed.2016.05.022. Epub 2016 May 27. Free Radic Biol Med. 2016. PMID: 27242268 Free PMC article. Review.
-
Warburg and Crabtree effects in premalignant Barrett's esophagus cell lines with active mitochondria.PLoS One. 2013;8(2):e56884. doi: 10.1371/journal.pone.0056884. Epub 2013 Feb 27. PLoS One. 2013. PMID: 23460817 Free PMC article.
-
Identification of a prognosis‑associated signature associated with energy metabolism in triple‑negative breast cancer.Oncol Rep. 2020 Sep;44(3):819-837. doi: 10.3892/or.2020.7657. Epub 2020 Jun 23. Oncol Rep. 2020. PMID: 32582991 Free PMC article.
References
-
- Whitaker-Menezes D, Martinez-Outschoorn UE, Lin Z, Ertel A, Flomenberg N, Witkiewicz AK, et al. Evidence for a stromal-epithelial “lactate shuttle” in human tumors: MCT4 is a marker of oxidative stress in cancer-associated fibroblasts. Cell Cycle. 2011;10:1772–1783. doi: 10.4161/cc.10.11.15659. - DOI - PMC - PubMed
-
- Martinez-Outschoorn UE, Pavlides S, Howell A, Pestell RG, Tanowitz HB, Sotgia F, et al. Stromalepithelial metabolic coupling in cancer: integrating autophagy and metabolism in the tumor microenvironment. Int J Biochem Cell Biol. 2011;43:1045–1051. doi: 10.1016/j.biocel.2011.01.023. - DOI - PMC - PubMed
-
- Pavlides S, Vera I, Gandara R, Sneddon S, Pestell R, Mercier I, et al. Warburg Meets Autophagy: Cancer Associated Fibroblasts Accelerate Tumor Growth and Metastasis Via Oxidative Stress, Mitophagy and Aerobic Glycolysis. Antioxid Redox Signal. 2011 doi: 10.1089/ars.2011.4243. In press. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 CA075503/CA/NCI NIH HHS/United States
- R01 CA098779/CA/NCI NIH HHS/United States
- R01-CA-120876/CA/NCI NIH HHS/United States
- R01 CA120876/CA/NCI NIH HHS/United States
- R01-CA-70896/CA/NCI NIH HHS/United States
- R01-CA-098779/CA/NCI NIH HHS/United States
- R01-CA-86072/CA/NCI NIH HHS/United States
- R01-AR-055660/AR/NIAMS NIH HHS/United States
- R01-CA-080250/CA/NCI NIH HHS/United States
- R01 CA070896/CA/NCI NIH HHS/United States
- R01 CA107382/CA/NCI NIH HHS/United States
- P30 CA056036/CA/NCI NIH HHS/United States
- P30-CA-56036/CA/NCI NIH HHS/United States
- R01-CA-107382/CA/NCI NIH HHS/United States
- R01 AR055660/AR/NIAMS NIH HHS/United States
- R01-CA-75503/CA/NCI NIH HHS/United States
- R01 CA080250/CA/NCI NIH HHS/United States
- R01 CA086072/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical