

Abcg2 expression marks tissue-specific stem cells in multiple organs in a mouse progeny tracking model
- PMID: 22134889
- PMCID: PMC4711397
- DOI: 10.1002/stem.1002
Abcg2 expression marks tissue-specific stem cells in multiple organs in a mouse progeny tracking model
Abstract
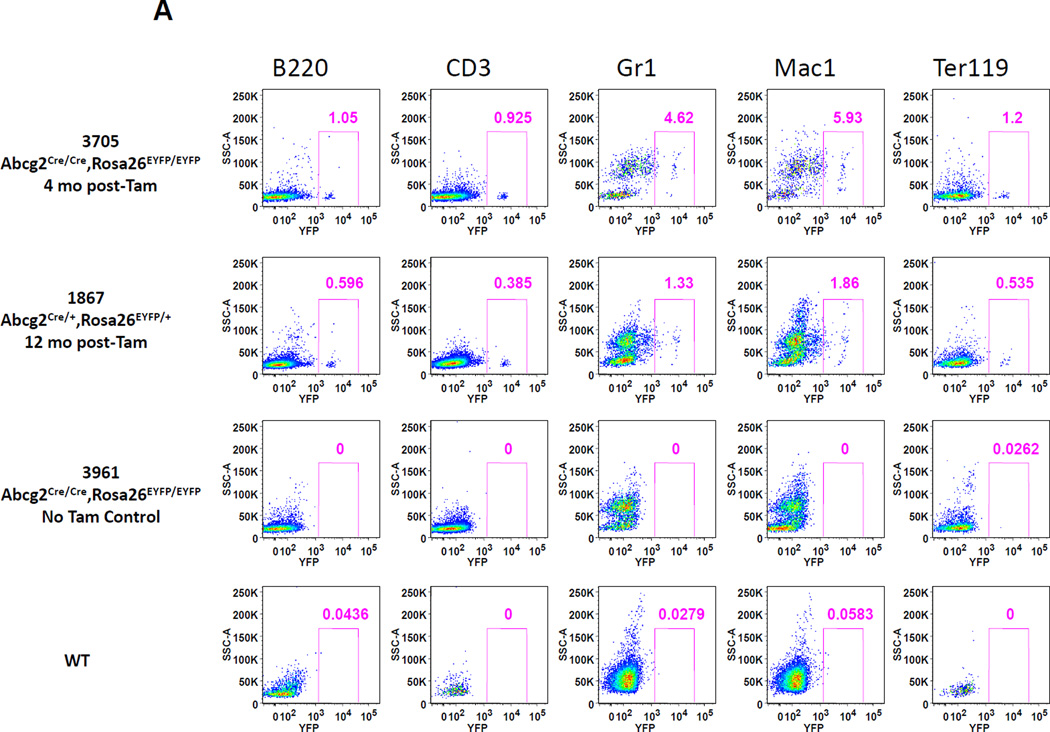
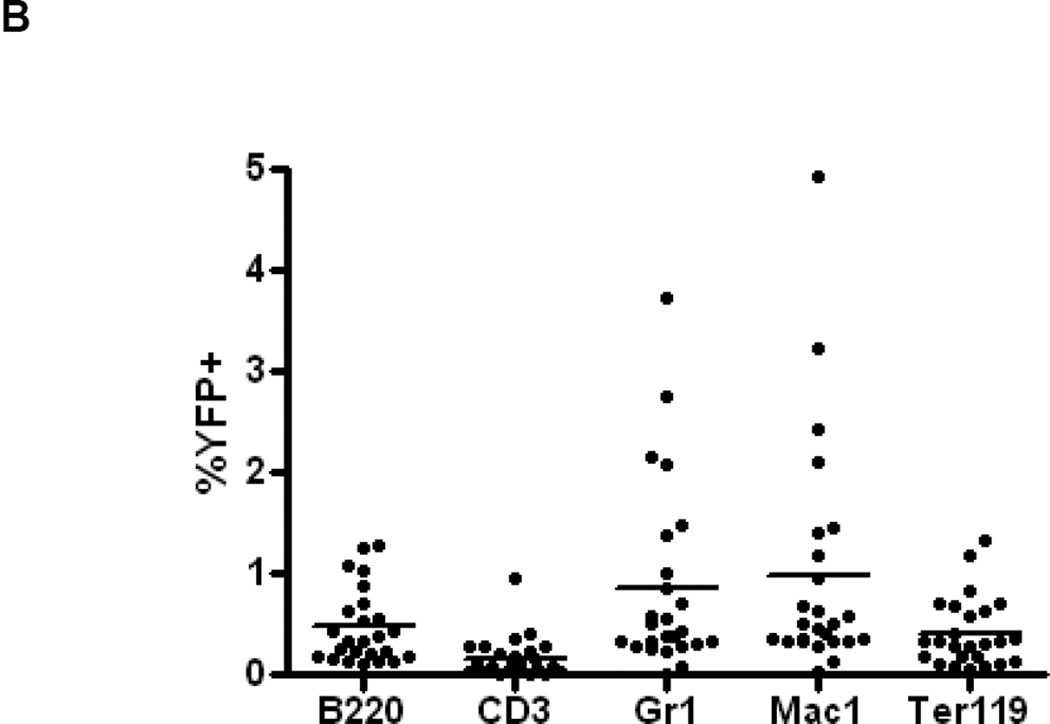
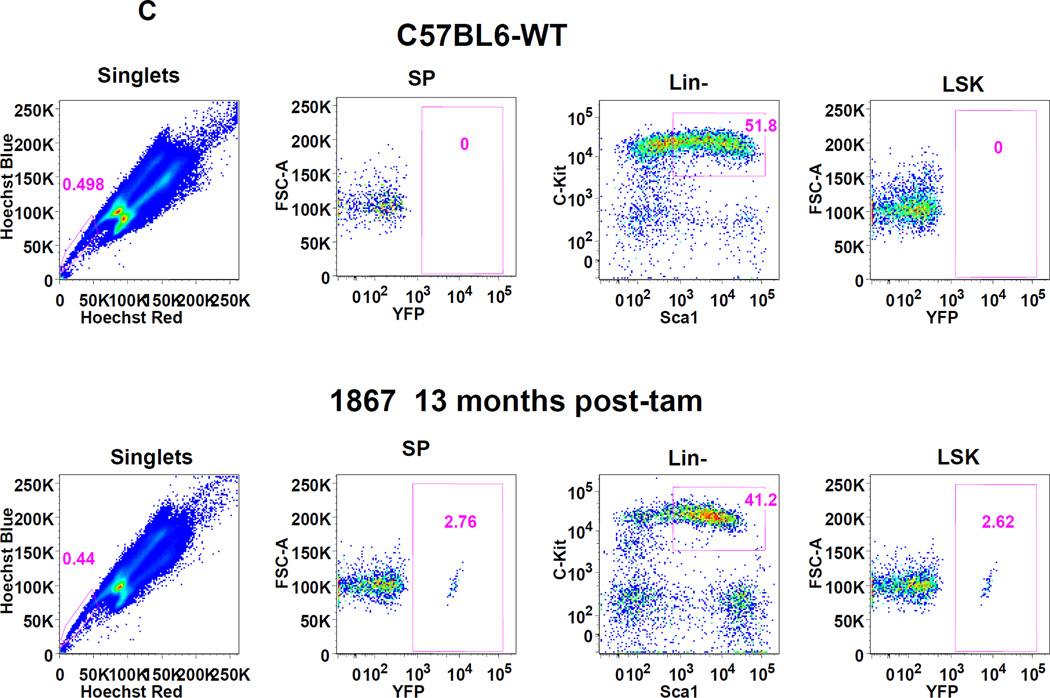
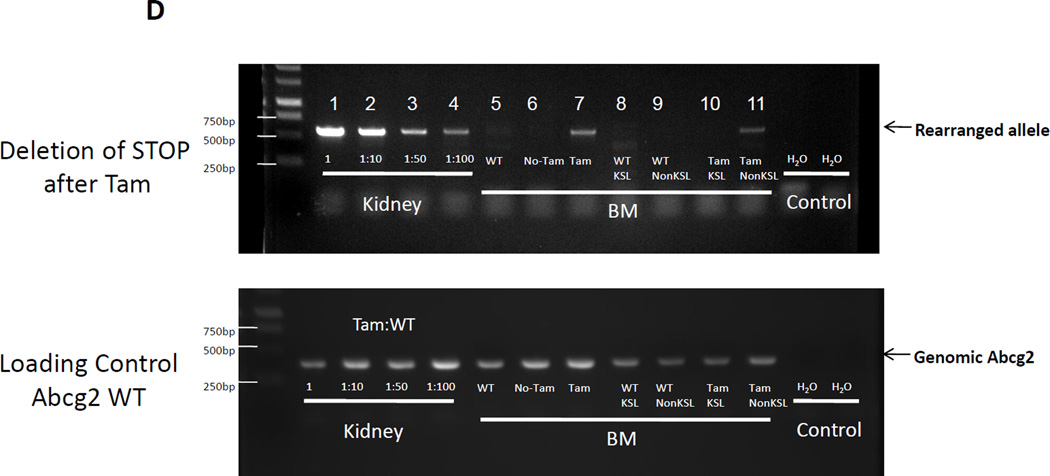
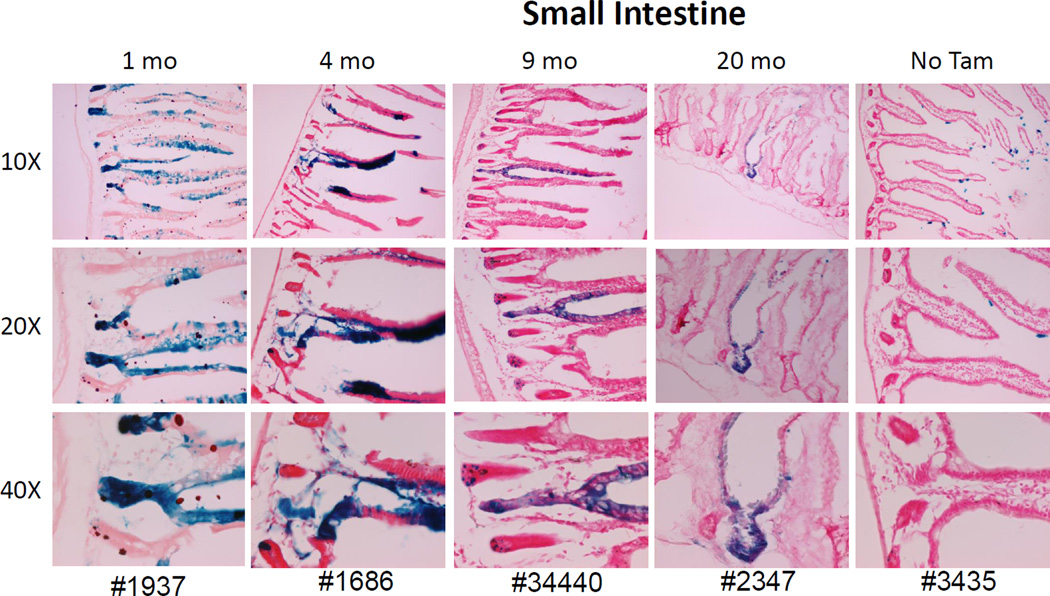
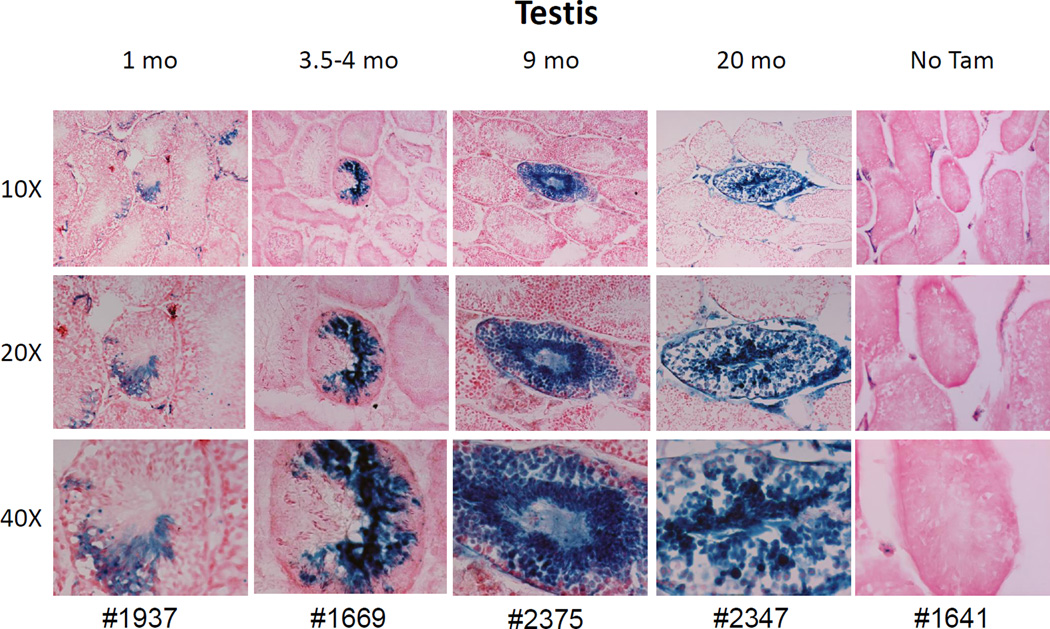
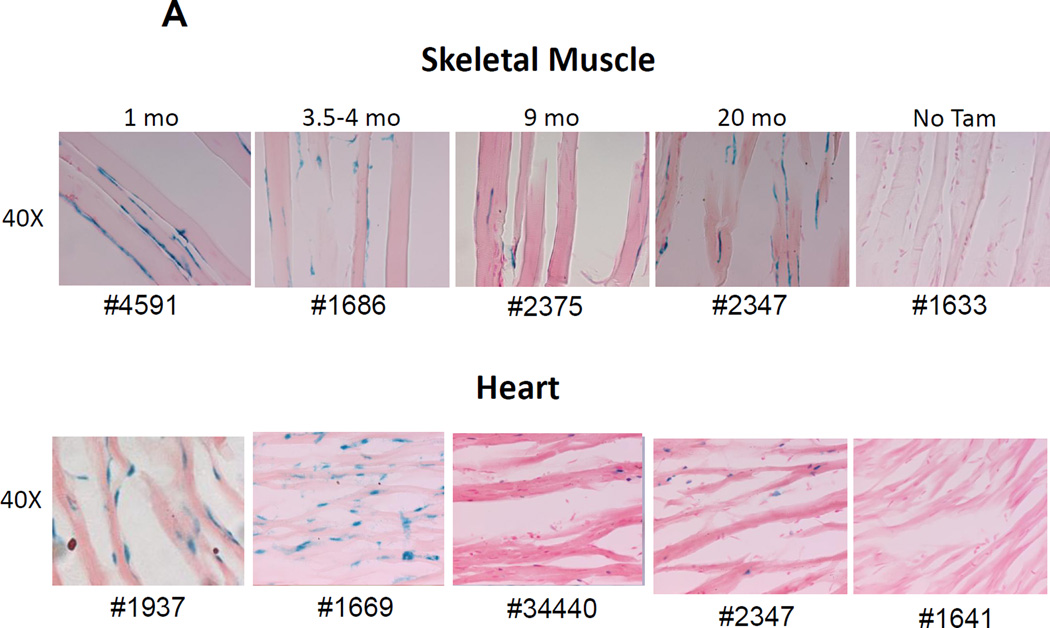
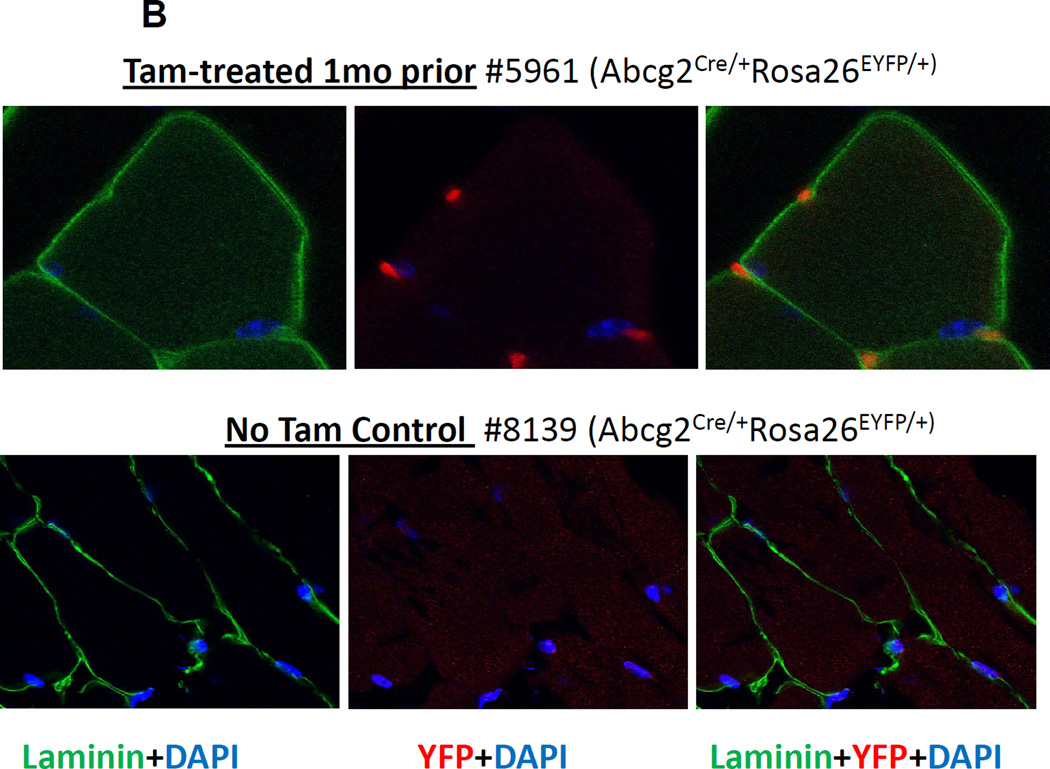
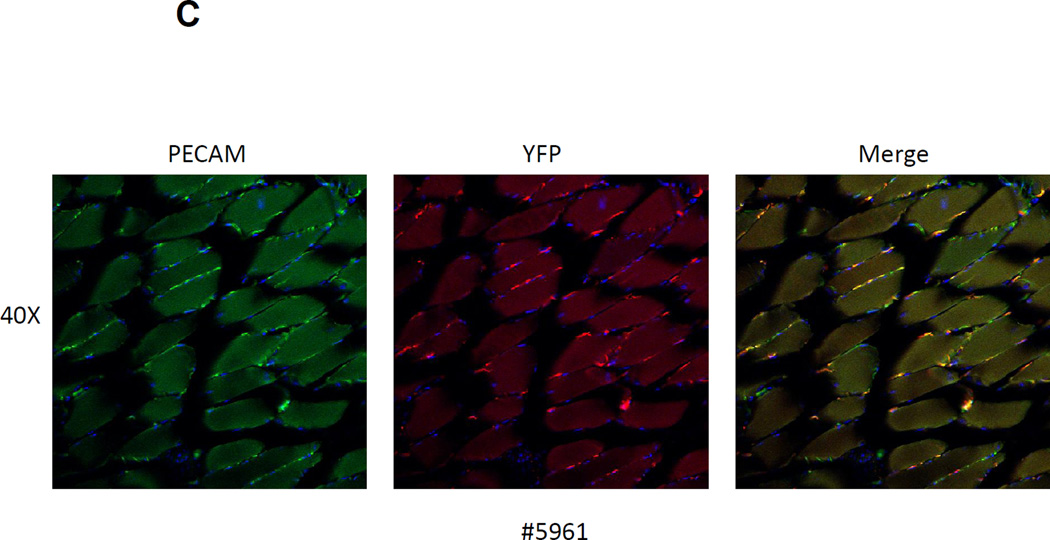
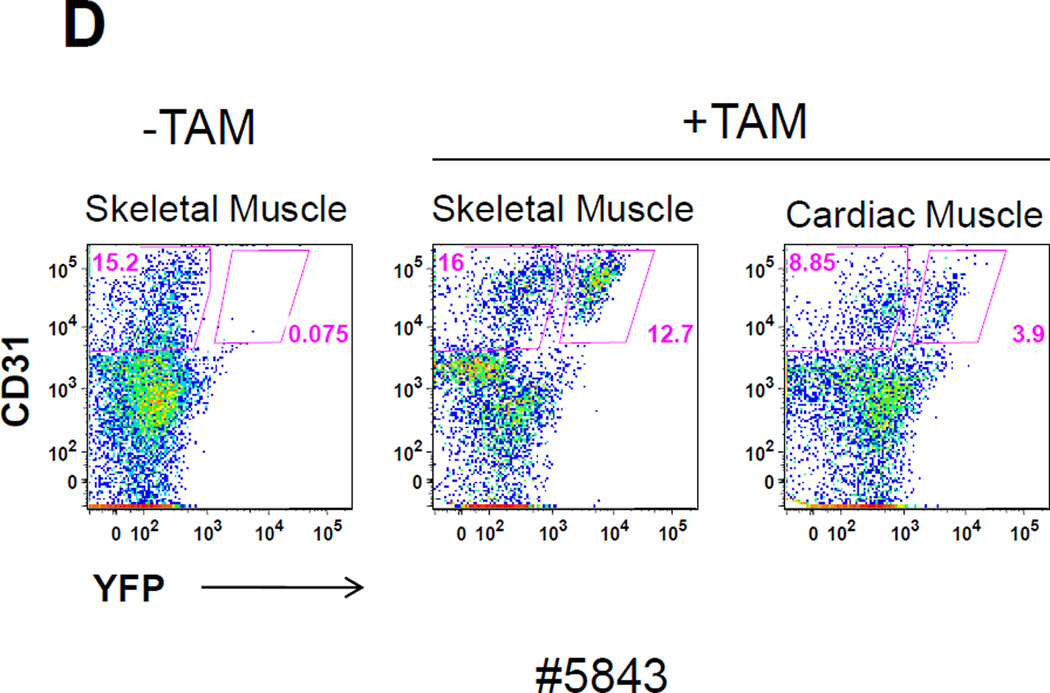
The side population phenotype is associated with the Hoechst dye efflux activity of the Abcg2 transporter and identifies hematopoietic stem cells (HSCs) in the bone marrow. This association suggests the direct use of Abcg2 expression to identify adult stem cells in various other organs. We have generated a lineage tracing mouse model based on an allele that coexpresses both Abcg2 and a CreERT2 expression cassette. By crossing these mice with lox-STOP-lox reporter lines (LacZ or YFP), cells that express Abcg2 and their progeny were identified following treatment with tamoxifen (Tam). In the liver and kidney, in which mature cells express Abcg2, reporter gene expression verified the expected physiologic expression pattern of the recombinant allele. Long-term marking of HSCs was seen in multiple peripheral blood lineages from adult mice, demonstrating that Abcg2(+) bone marrow HSCs contribute to steady-state hematopoiesis. Stem cell tracing patterns were seen in the small intestine and in seminiferous tubules in the testis 20 months after Tam treatment, proving that stem cells from these organs express Abcg2. Interstitial cells from skeletal and cardiac muscle were labeled, and some cells were costained with endothelial markers, raising the possibility that these cells may function in the repair response to muscle injury. Altogether, these studies prove that Abcg2 is a stem cell marker for blood, small intestine, testicular germ cells, and possibly for injured skeletal and/or cardiac muscle and provide a new model for studying stem cell activity that does not require transplant-based assays.
Copyright © 2011 AlphaMed Press.
Conflict of interest statement
Figures
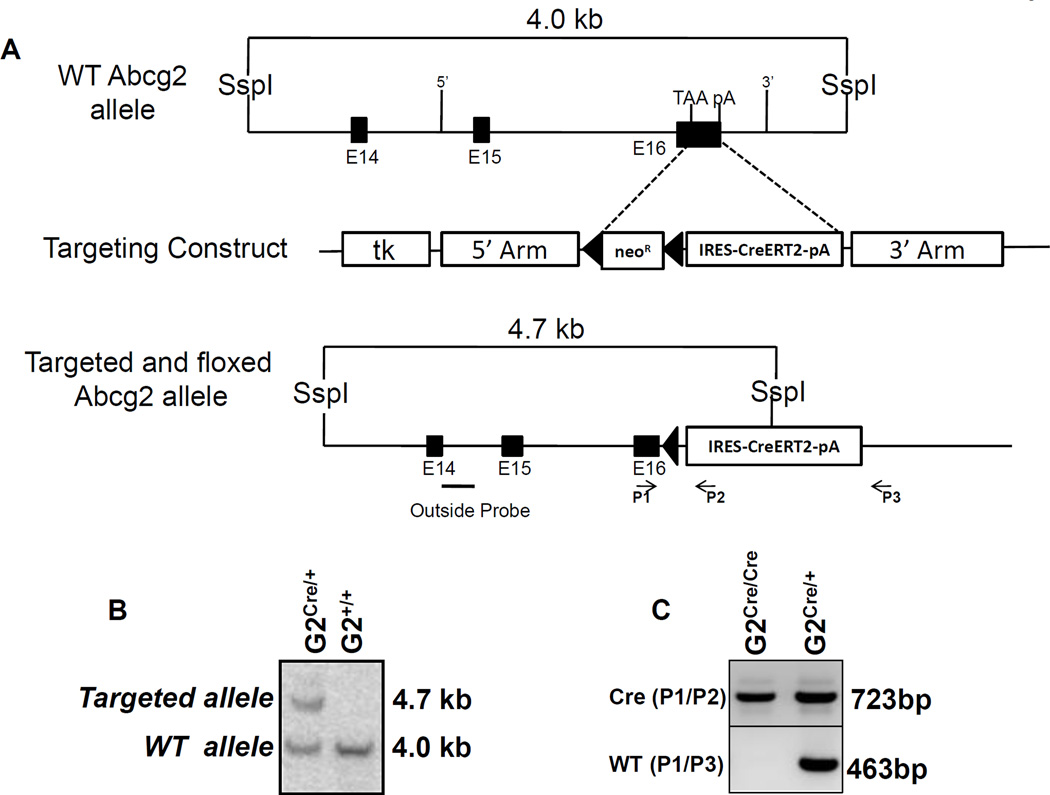
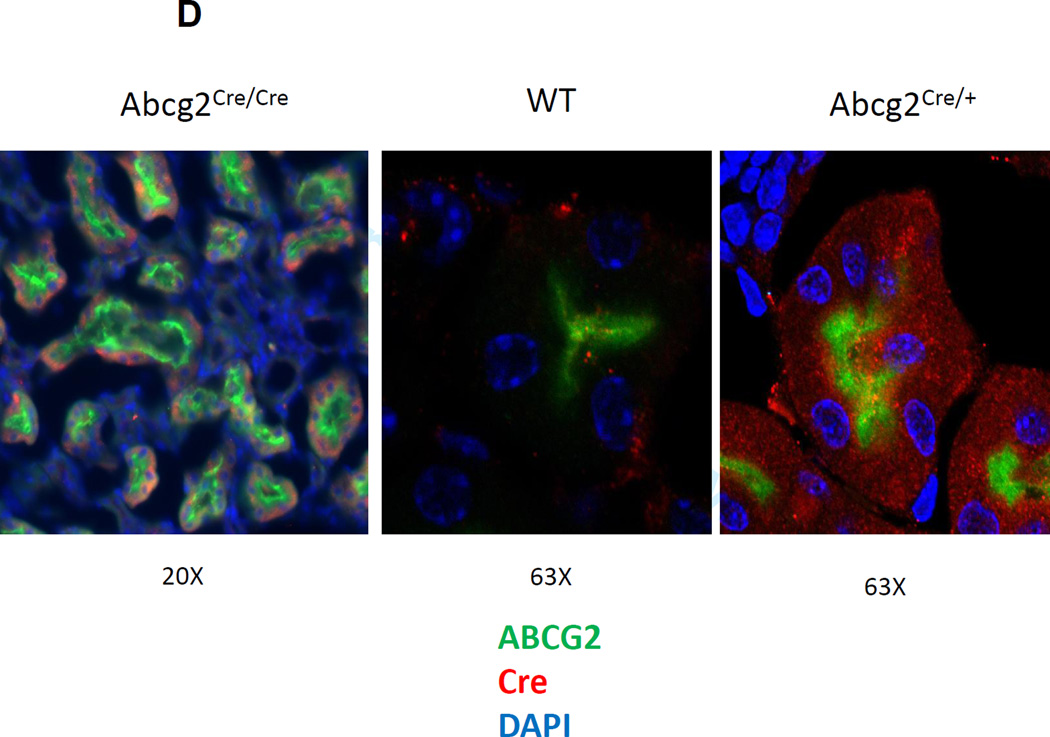
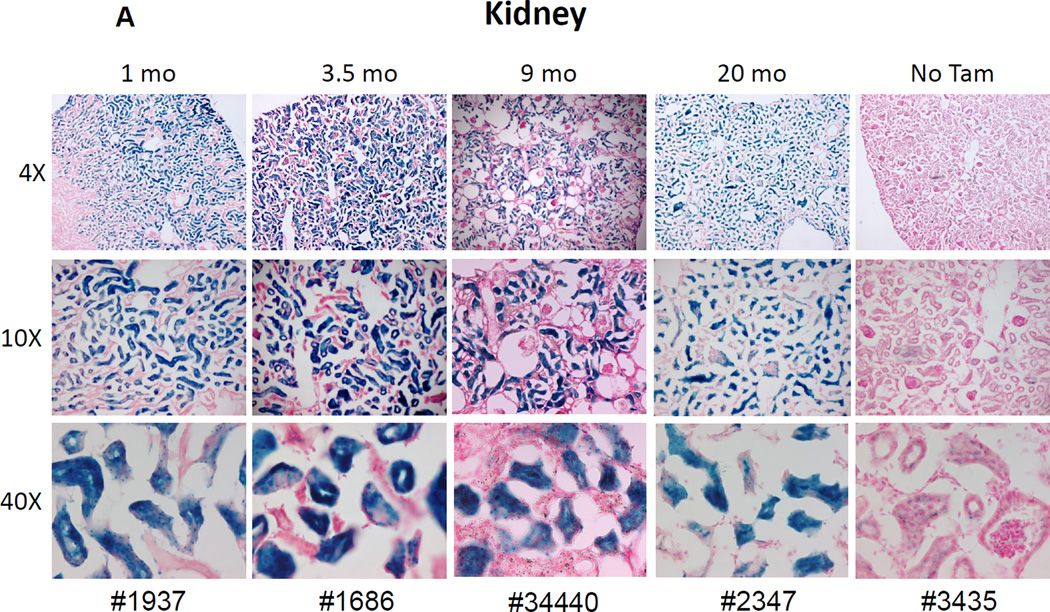
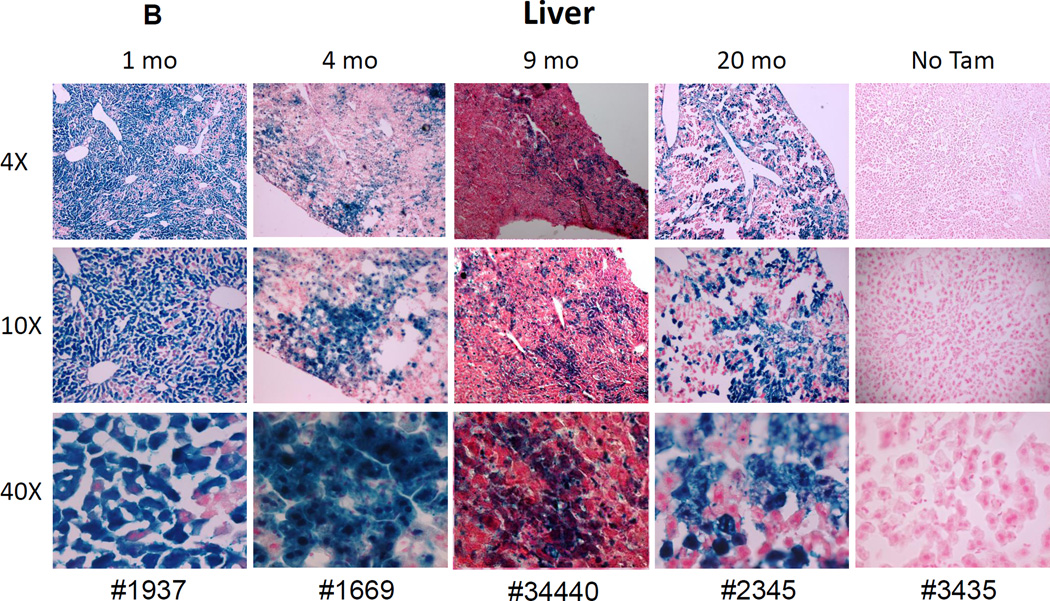
References
-
- Goodell MA, Rosenzweig M, Kim H, et al. Dye efflux studies suggest that hematopoietic stem cells expressing low or undetectable levels of CD34 antigen exist in multiple species. Nat. Med. 1997;3(12):1337–1345. - PubMed
-
- Zhou S, Zong Y, Lu T, et al. Hematopoietic cells from mice that are deficient in both Bcrp1/Abcg2 and Mdr1a/1b develop normally but are sensitized to mitoxantrone. Biotechniques. 2003;35(6):1248–1252. - PubMed
-
- Zhou S, Schuetz JD, Bunting KD, et al. The ABC transporter Bcrp1/ABCG2 is expressed in a wide variety of stem cells and is a molecular determinant of the side-population phenotype. Nat. Med. 2001;7(9):1028–1034. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
