

Characterization of heparin-induced glyceraldehyde-3-phosphate dehydrogenase early amyloid-like oligomers and their implication in α-synuclein aggregation
- PMID: 22134915
- PMCID: PMC3268401
- DOI: 10.1074/jbc.M111.303503
Characterization of heparin-induced glyceraldehyde-3-phosphate dehydrogenase early amyloid-like oligomers and their implication in α-synuclein aggregation
Abstract
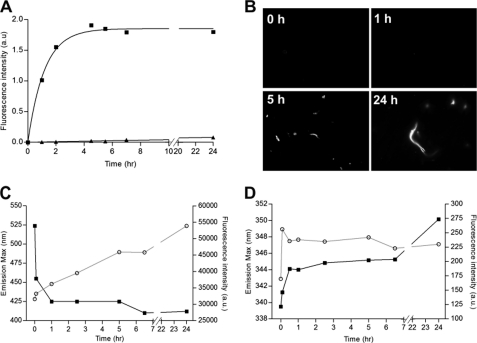
Lewy bodies and Lewy neurites, neuropathological hallmarks of several neurological diseases, are mainly made of filamentous assemblies of α-synuclein. However, other macromolecules including Tau, ubiquitin, glyceraldehyde-3-phosphate dehydrogenase, and glycosaminoglycans are routinely found associated with these amyloid deposits. Glyceraldehyde-3-phosphate dehydrogenase is a glycolytic enzyme that can form fibrillar aggregates in the presence of acidic membranes, but its role in Parkinson disease is still unknown. In this work, the ability of heparin to trigger the amyloid aggregation of this protein at physiological conditions of pH and temperature is demonstrated by infrared and fluorescence spectroscopy, dynamic light scattering, small angle x-ray scattering, circular dichroism, and fluorescence microscopy. Aggregation proceeds through the formation of short rod-like oligomers, which elongates in one dimension. Heparan sulfate was also capable of inducing glyceraldehyde-3-phosphate dehydrogenase aggregation, but chondroitin sulfates A, B, and C together with dextran sulfate had a negligible effect. Aided with molecular docking simulations, a putative binding site on the protein is proposed providing a rational explanation for the structural specificity of heparin and heparan sulfate. Finally, it is demonstrated that in vitro the early oligomers present in the glyceraldehyde-3-phosphate dehydrogenase fibrillation pathway promote α-synuclein aggregation. Taking into account the toxicity of α-synuclein prefibrillar species, the heparin-induced glyceraldehyde-3-phosphate dehydrogenase early oligomers might come in useful as a novel therapeutic strategy in Parkinson disease and other synucleinopathies.
Figures







References
-
- Tong J., Wong H., Guttman M., Ang L. C., Forno L. S., Shimadzu M., Rajput A. H., Muenter M. D., Kish S. J., Hornykiewicz O., Furukawa Y. (2010) Brain α-synuclein accumulation in multiple system atrophy, Parkinson's disease and progressive supranuclear palsy. A comparative investigation. Brain 133, 172–188 - PubMed
-
- Nussbaum R. L., Ellis C. E. (2003) Alzheimer's disease and Parkinson's disease. N. Engl. J. Med. 348, 1356–1364 - PubMed
-
- Anderson J. P., Walker D. E., Goldstein J. M., de Laat R., Banducci K., Caccavello R. J., Barbour R., Huang J., Kling K., Lee M., Diep L., Keim P. S., Shen X., Chataway T., Schlossmacher M. G., Seubert P., Schenk D., Sinha S., Gai W. P., Chilcote T. J. (2006) Phosphorylation of Ser-129 is the dominant pathological modification of α-synuclein in familial and sporadic Lewy body disease. J. Biol. Chem. 281, 29739–29752 - PubMed
-
- Spillantini M. G., Schmidt M. L., Lee V. M., Trojanowski J. Q., Jakes R., Goedert M. (1997) α-Synuclein in Lewy bodies. Nature 388, 839–840 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
