

Review
doi: 10.1186/1741-7007-9-85.
Putting the pH into phosphatidic acid signaling
Affiliations
- PMID: 22136116
- PMCID: PMC3229452
- DOI: 10.1186/1741-7007-9-85
Item in Clipboard
Review
Putting the pH into phosphatidic acid signaling
BMC Biol.
.
Abstract
The lipid phosphatidic acid (PA) has important roles in cell signaling and metabolic regulation in all organisms. New evidence indicates that PA also has an unprecedented role as a pH biosensor, coupling changes in pH to intracellular signaling pathways. pH sensing is a property of the phosphomonoester headgroup of PA. A number of other potent signaling lipids also contain headgroups with phosphomonoesters, implying that pH sensing by lipids may be widespread in biology.
Figures
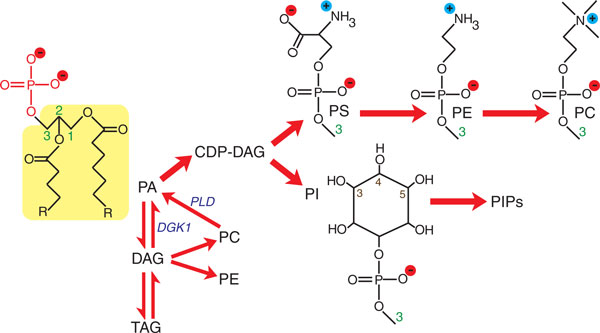
Phosphatidic acid is a key precursor in lipid metabolism. A simplified outline of the major lipids originating from phosphatidic acid (PA) is shown. These include glycerophospholipids: CDP-diacylglycerol (CDP-DAG), phosphatidylserine (PS), phosphatidylethanolamine (PE), phosphatidylcholine (PC), phosphatidylinositol (PI), and phosphoinositol phosphates (PIPs); and neutral lipids: diacylglycerol (DAG) and triacylglycerol (TAG). PA is drawn in its deprotonated form carrying a charge of 2-. Its structure is composed of a phosphomonoester headgroup (shown in red) attached to a DAG backbone (highlighted in yellow). This DAG backbone is composed of glycerol with two acyl chains attached at its sn1 and sn2 positions (green numerals indicate the sn positions of glycerol, R represents the remaining structure of each acyl chain that is not shown). All glycerophospholipids have their headgroups attached to the DAG backbone at the sn3 position of glycerol. The headgroups of PS, PE, PC and PI are shown for comparison and the positions of phosphorylation of the inositol ring in PIPs are labeled (3,4,5). The actions of DAG kinase (DGK) and phospholipase D (PLD) in regulating PA levels are also indicated (shown in blue italics).

Lipid signaling is concentration dependent. Lipid effector proteins are unable to bind membranes when the concentration of their target lipid is low (left). Cellular signals lead to the activation of lipid-modifying enzymes, which generate target lipids that can then recruit the effector proteins to the membrane (right). Conversely, other cellular signals can activate enzymes that convert the target lipid back to its original form or to another lipid, causing release of the effector from the membrane. In this example, the target lipid highlighted in yellow is also cone-shaped (for example, PA, PE or DAG) in contrast to the majority of membrane lipids, which are cylindrical in shape (for example, PC and PS; headgroups colored brown). The conical shape reduces packing of lipid headgroups, which exposes the hydrophobic acyl layer surrounding the target lipid. This may facilitate insertion of hydrophobic amino acids and effector binding.

Hydrogen bonding increases the charge of phosphatidic acid (PA) and lysophosphatidic acid (LPA). (a) The phosphomonoester headgroup of LPA forms an intramolecular hydrogen bond. In the protonated phosphomonoester of LPA (left) a proton is shared between two hydroxyl oxygens (purple dashed lines). Hydrogen bonding (green dashed lines) between the sn2 hydroxyl and the phosphomonoester of LPA competes with the shared proton for oxygen electrons, which facilitates dissociation of the proton lowering the pKa. (b) Hydrogen bonding between the phosphomonoester of PA and the primary amine of the headgroup of PE, or lysines and arginines of proteins, results in deprotonation of the phosphomonoester, lowering its pKa and increasing its negative charge. The abundance of PE in cellular membranes is likely a significant factor regulating the pKa of both PA and LPA [30]. Hydrogen bonding between proteins and PA is described by the electrostatic/hydrogen bond switch mechanism [29].
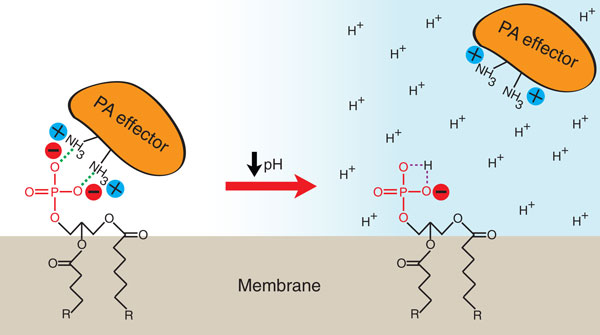
Phosphatidic acid is a pH biosensor. Effector proteins bind PA according to the electrostatic/hydrogen-bond switch mechanism [29]. Electrostatic interactions and hydrogen bonding (green dashed lines) between the primary amines of basic amino acids in the effector and the deprotonated phosphomonoester of PA are shown. A decrease in intracellular pH below the pKa of PA results in increased protonation of its phosphomonoester, which reduces the strength of electrostatic interactions with the effector, resulting in its dissociation from the membrane.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
