

miR-124 acts through CoREST to control onset of Sema3A sensitivity in navigating retinal growth cones
- PMID: 22138647
- PMCID: PMC3661270
- DOI: 10.1038/nn.2979
miR-124 acts through CoREST to control onset of Sema3A sensitivity in navigating retinal growth cones
Abstract
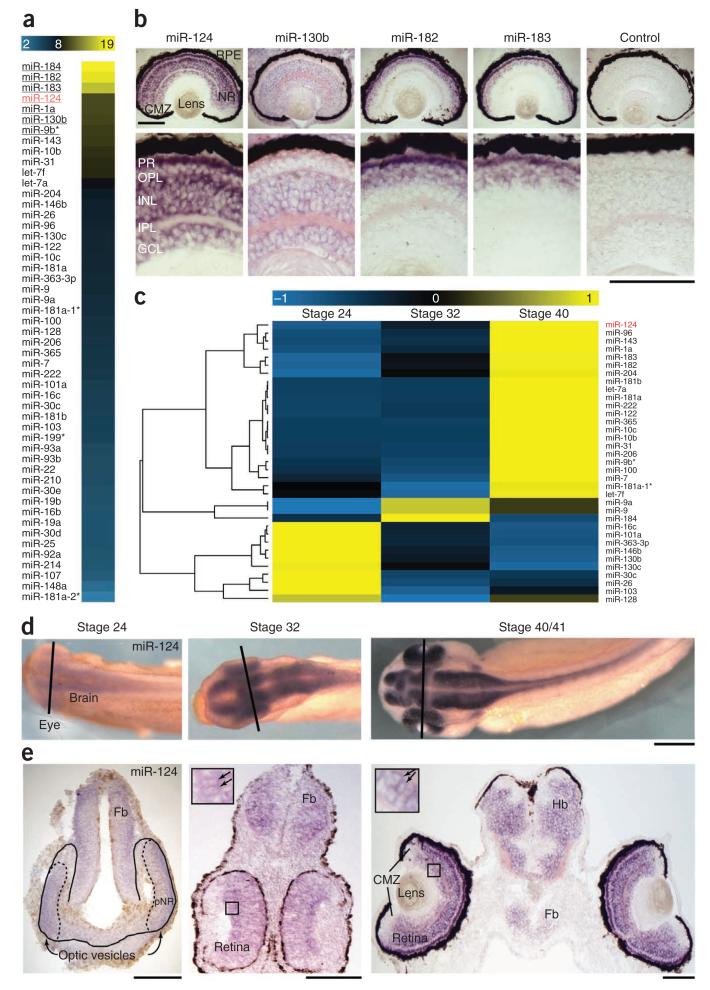
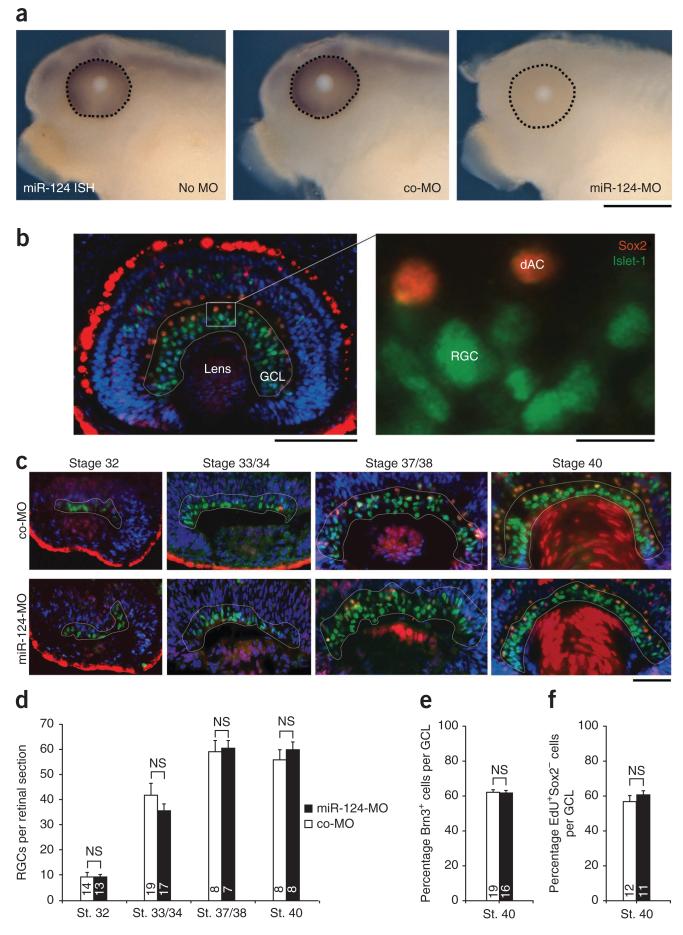
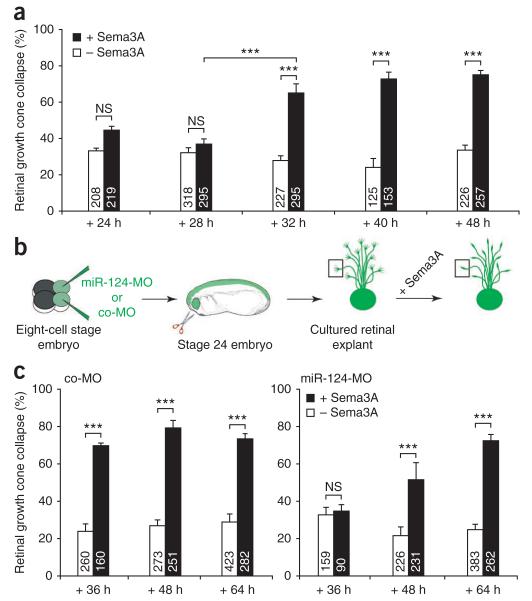
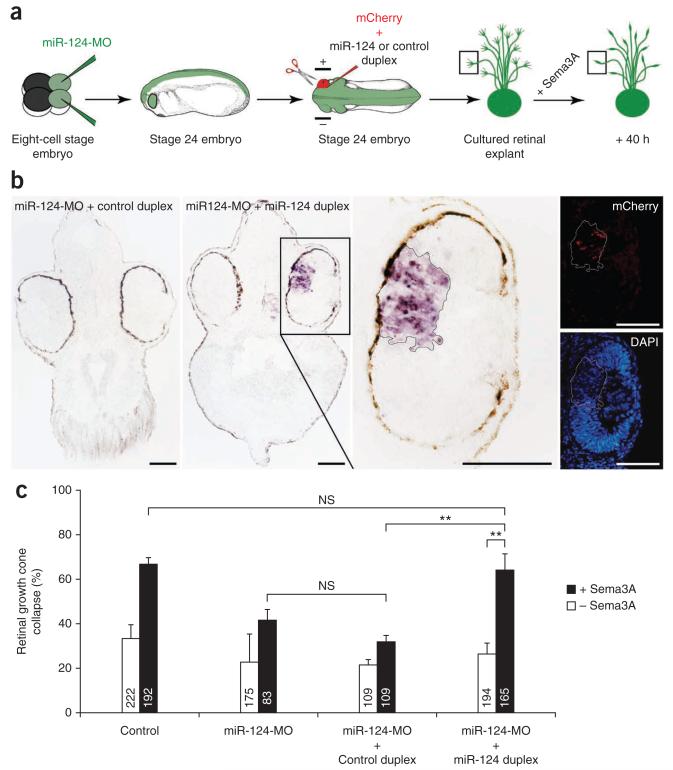
During axon pathfinding, growth cones commonly show changes in sensitivity to guidance cues that follow a cell-intrinsic timetable. The cellular timer mechanisms that regulate such changes are, however, poorly understood. Here we have investigated microRNAs (miRNAs) in the timing control of sensitivity to the semaphorin Sema3A in Xenopus laevis retinal ganglion cell (RGC) growth cones. A developmental profiling screen identified miR-124 as a candidate timer. Loss of miR-124 delayed the onset of Sema3A sensitivity and concomitant neuropilin-1 (NRP1) receptor expression and caused cell-autonomous pathfinding errors. CoREST, a cofactor of a NRP1 repressor, was newly identified as a target and mediator of miR-124 for this highly specific temporal aspect of RGC growth cone responsiveness. Our findings indicate that miR-124 is important in regulating the intrinsic temporal changes in RGC growth cone sensitivity and suggest that miRNAs may act broadly as linear timers in vertebrate neuronal development.
Figures
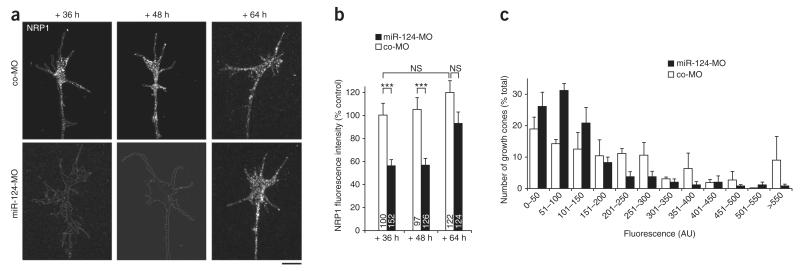
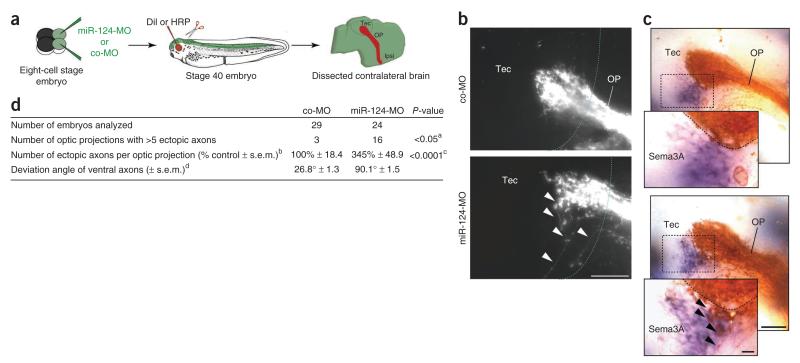
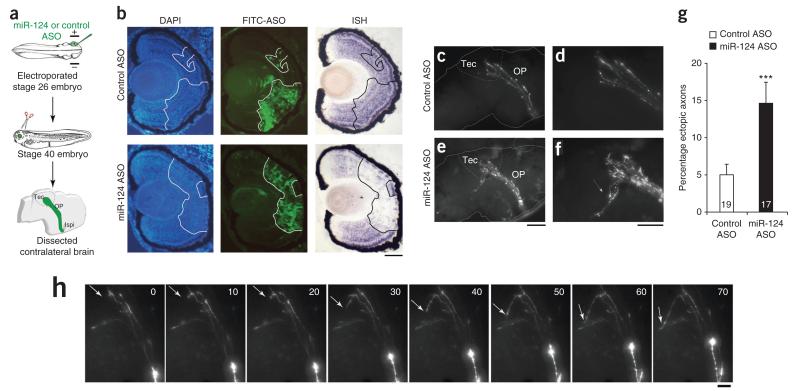
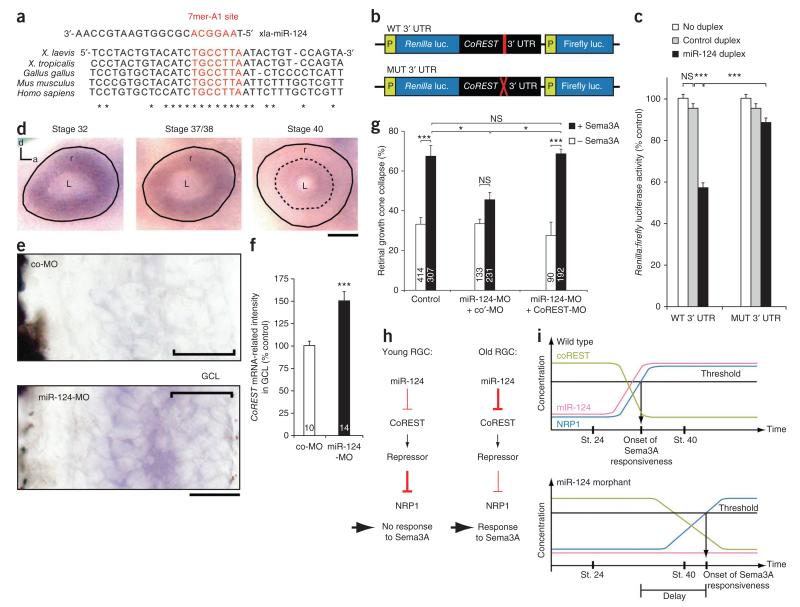
References
-
- Tessier-Lavigne M, Goodman CS. The molecular biology of axon guidance. Science. 1996;274:1123–1133. - PubMed
-
- Dickson BJ. Molecular mechanisms of axon guidance. Science. 2002;298:1959–1964. - PubMed
-
- Zou Y, Stoeckli E, Chen H, Tessier-Lavigne M. Squeezing axons out of the gray matter: a role for slit and semaphorin proteins from midline and ventral spinal cord. Cell. 2000;102:363–375. - PubMed
-
- Stein E, Tessier-Lavigne M. Hierarchical organization of guidance receptors: silencing of netrin attraction by slit through a Robo/DCC receptor complex. Science. 2001;291:1928–1938. - PubMed
-
- Shewan D, Dwivedy A, Anderson R, Holt CE. Age-related changes underlie switch in netrin-1 responsiveness as growth cones advance along visual pathway. Nat. Neurosci. 2002;5:955–962. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
