

Ndc10 is a platform for inner kinetochore assembly in budding yeast
- PMID: 22139014
- PMCID: PMC3252399
- DOI: 10.1038/nsmb.2178
Ndc10 is a platform for inner kinetochore assembly in budding yeast
Abstract
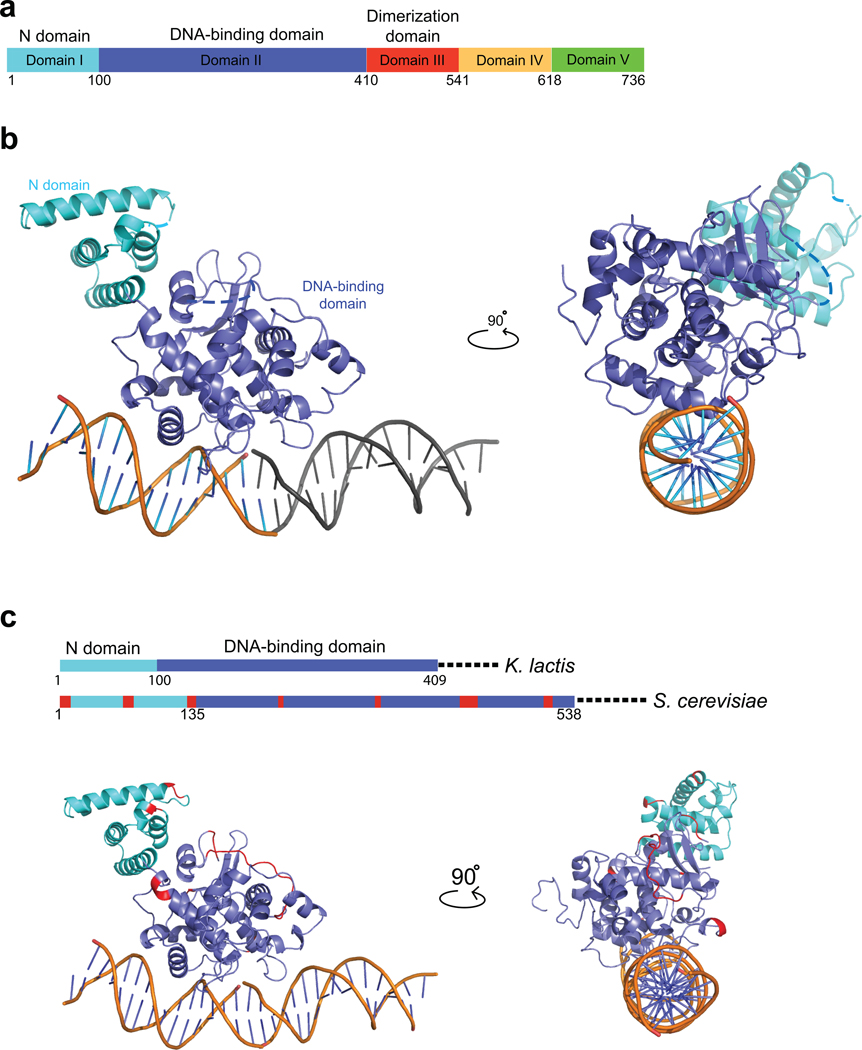
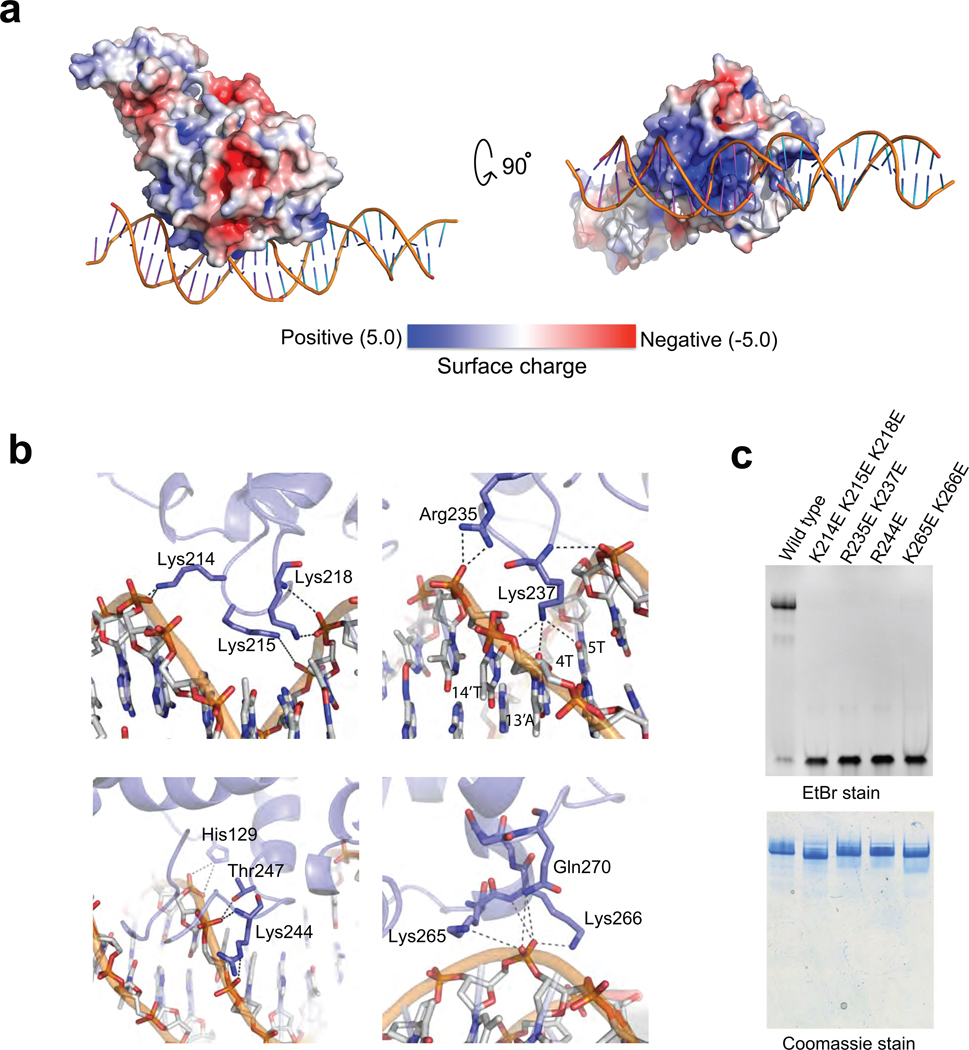
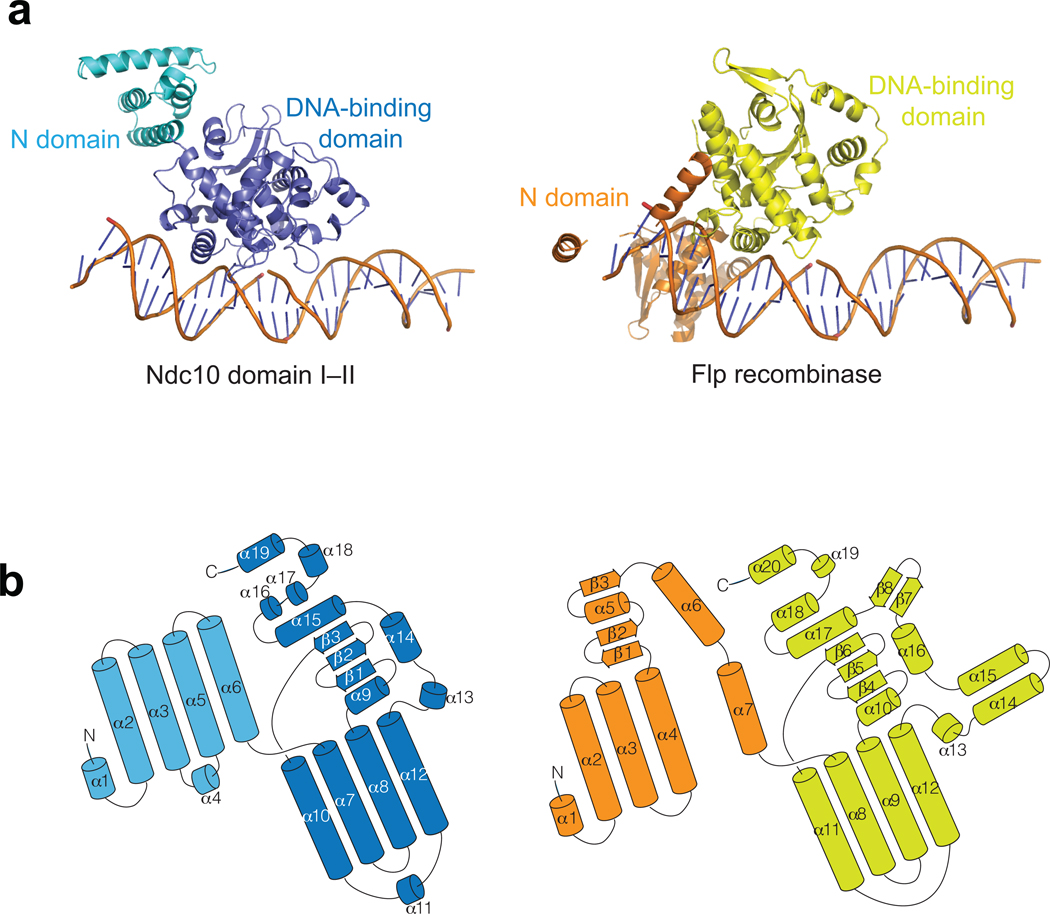
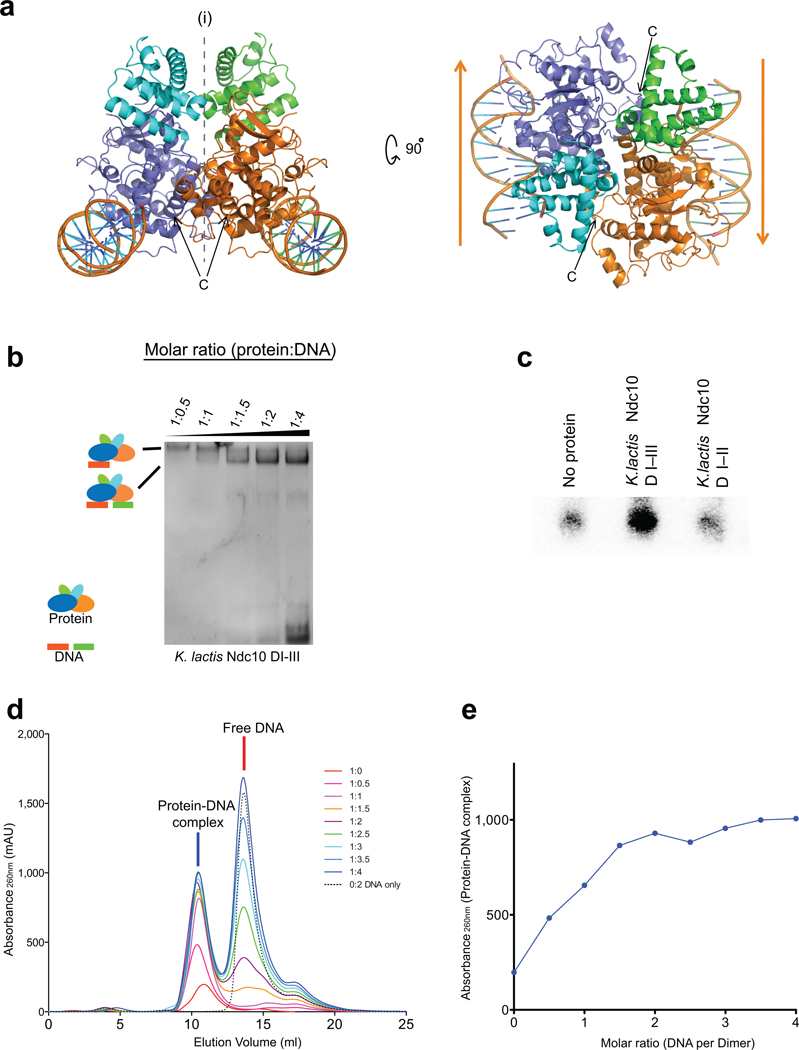
Kinetochores link centromeric DNA to spindle microtubules and ensure faithful chromosome segregation during mitosis. In point-centromere yeasts, the CBF3 complex Skp1-Ctf13-(Cep3)(2)-(Ndc10)(2) recognizes a conserved centromeric DNA element through contacts made by Cep3 and Ndc10. We describe here the five-domain organization of Kluyveromyces lactis Ndc10 and the structure at 2.8 Å resolution of domains I-II (residues 1-402) bound to DNA. The structure resembles tyrosine DNA recombinases, although it lacks both endonuclease and ligase activities. Structural and biochemical data demonstrate that each subunit of the Ndc10 dimer binds a separate fragment of DNA, suggesting that Ndc10 stabilizes a DNA loop at the centromere. We describe in vitro association experiments showing that specific domains of Ndc10 interact with each of the known inner-kinetochore proteins or protein complexes in budding yeast. We propose that Ndc10 provides a central platform for inner-kinetochore assembly.
Figures
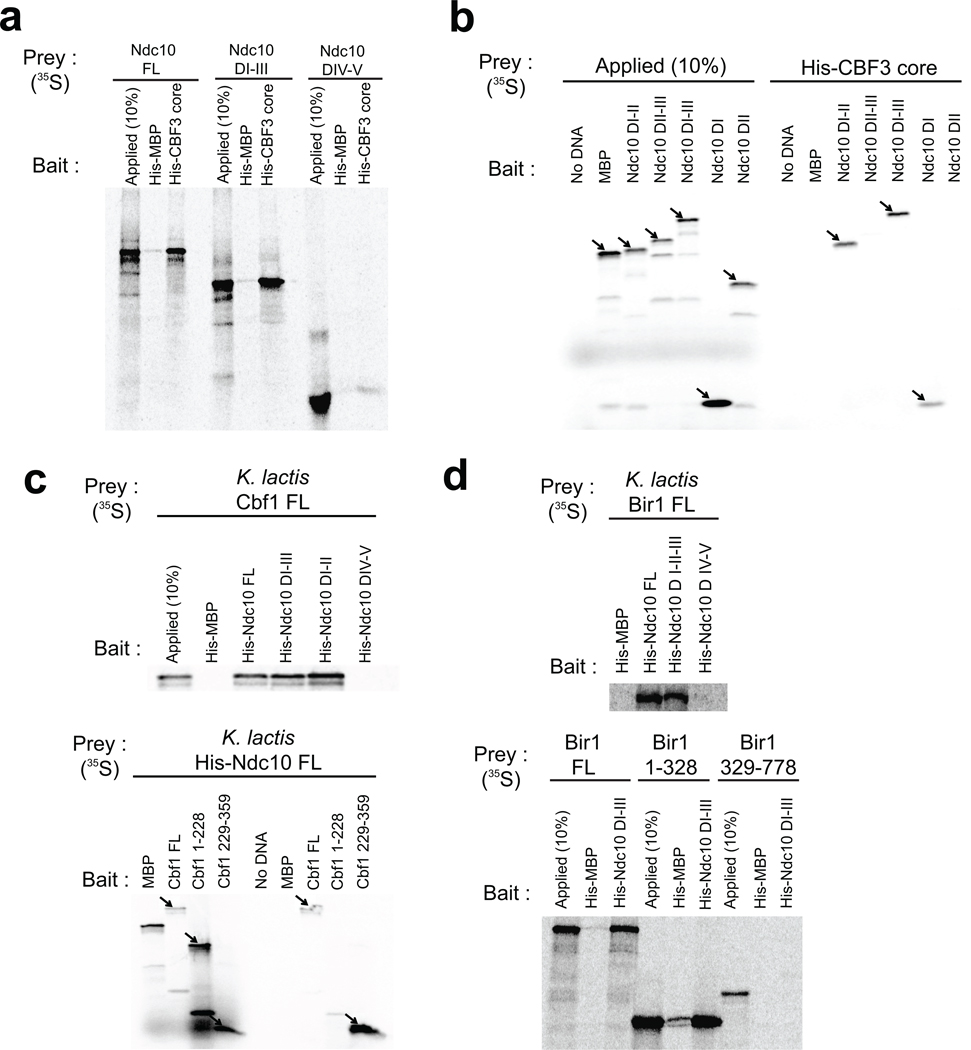
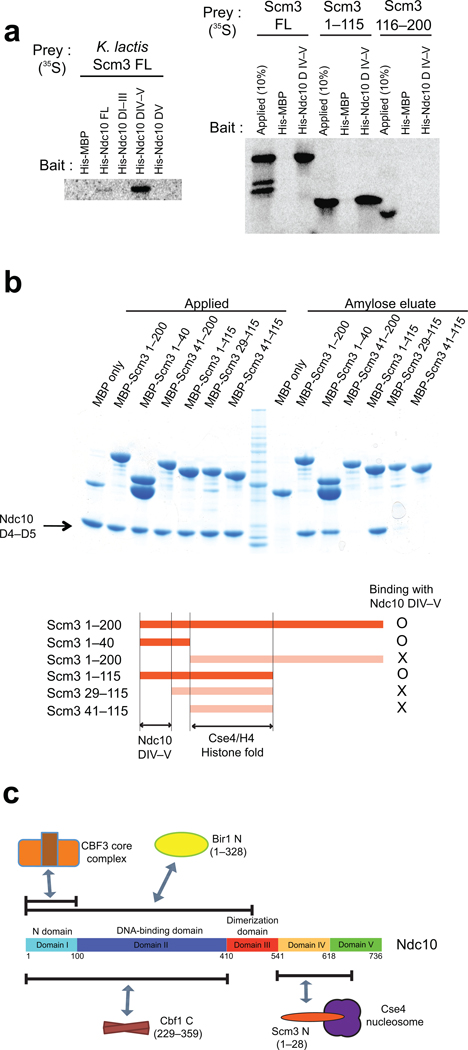
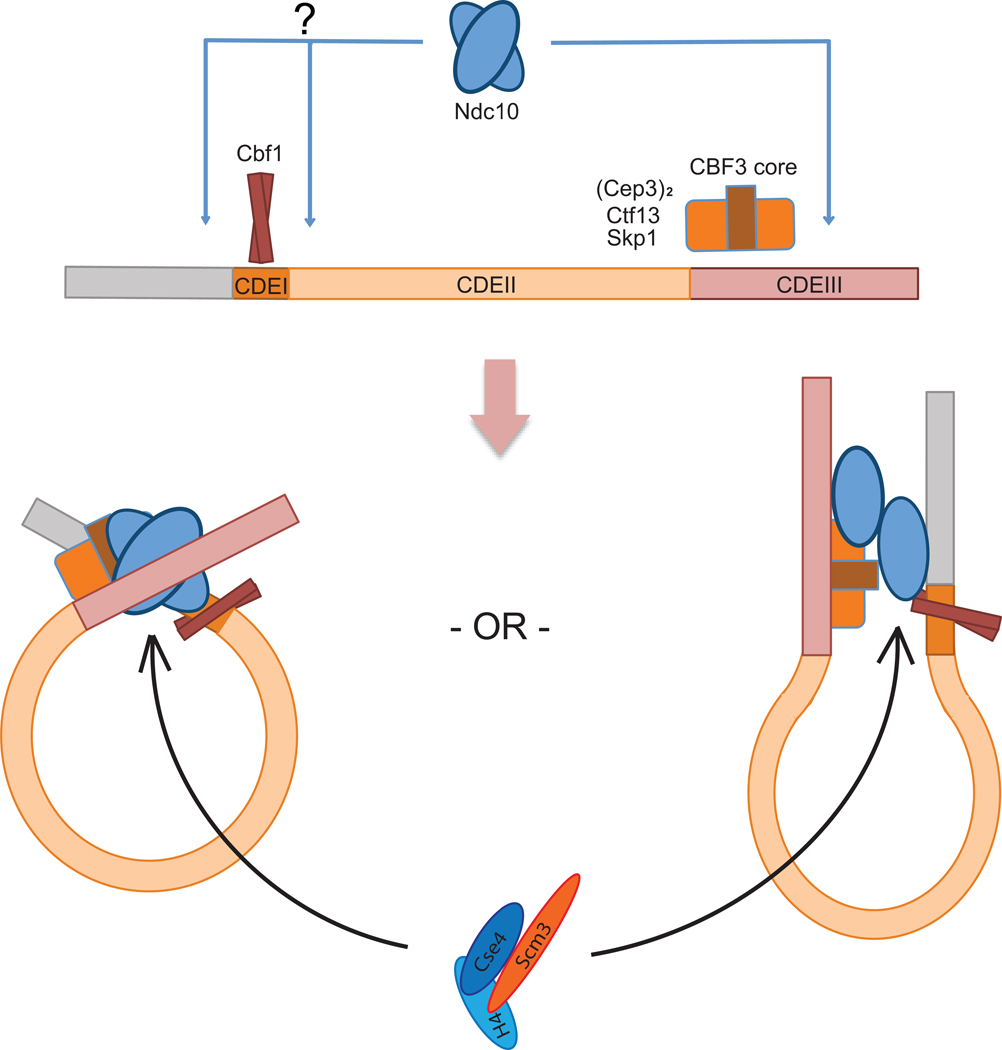
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
