

Neural representations of faces and limbs neighbor in human high-level visual cortex: evidence for a new organization principle
- PMID: 22139022
- PMCID: PMC3535411
- DOI: 10.1007/s00426-011-0392-x
Neural representations of faces and limbs neighbor in human high-level visual cortex: evidence for a new organization principle
Abstract
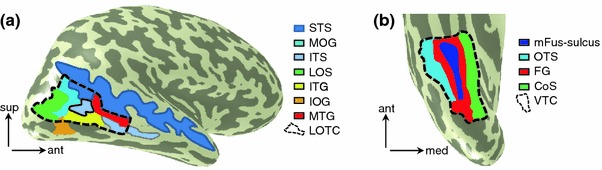
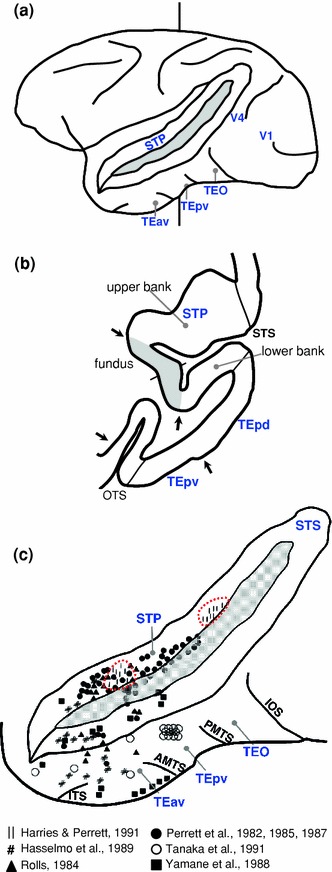
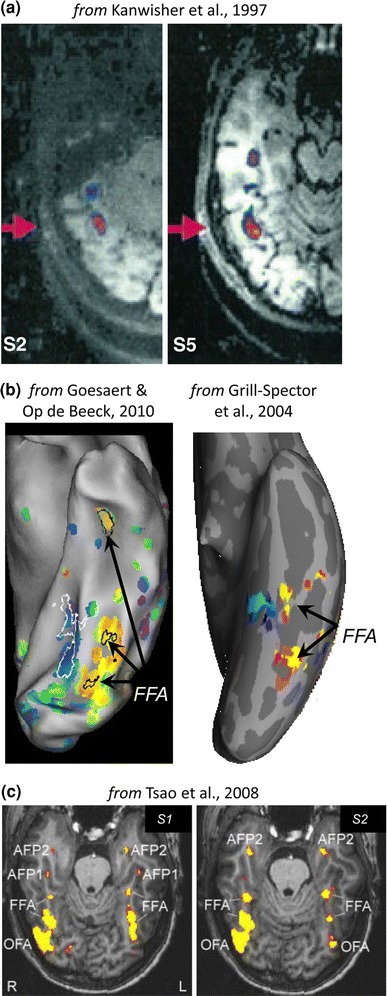
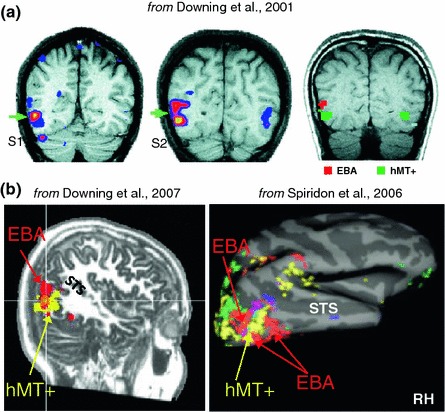
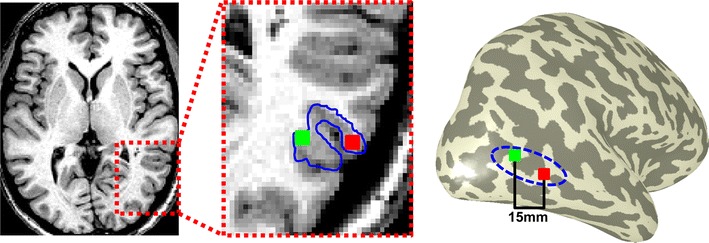
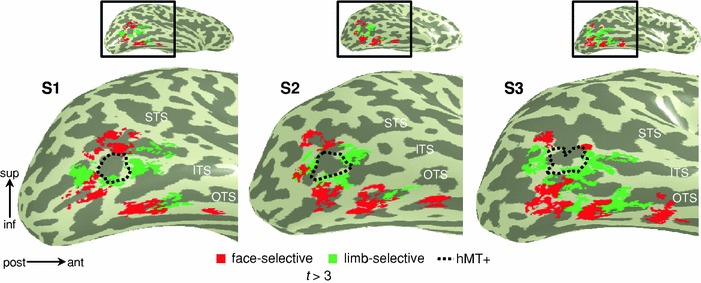
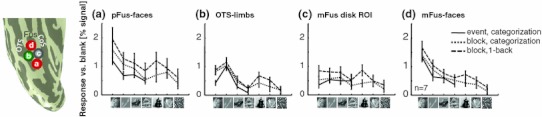
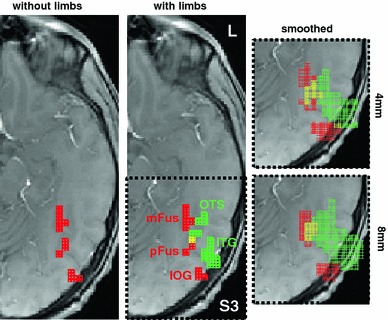
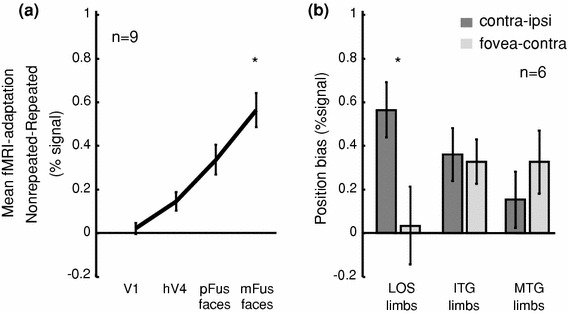
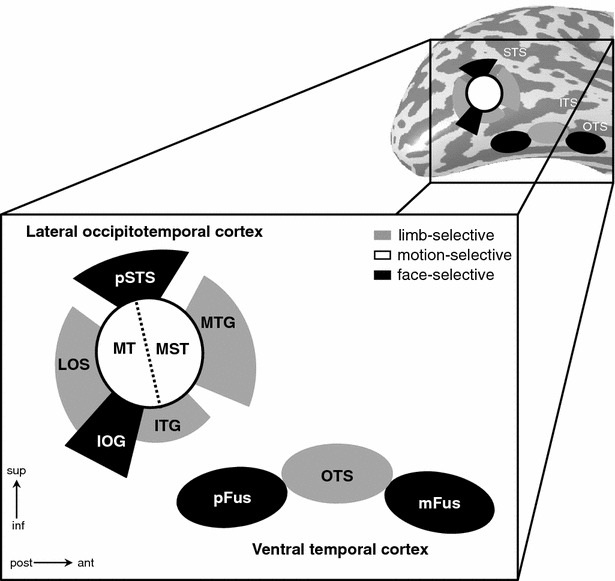
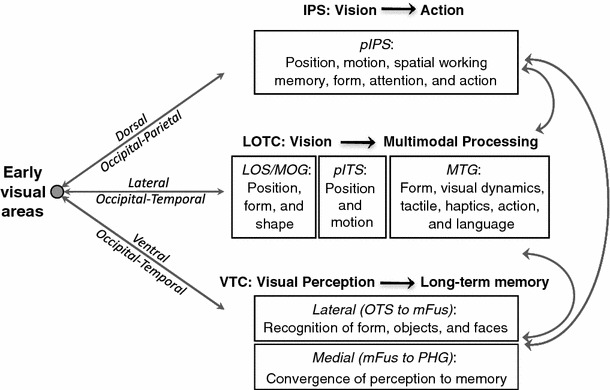
Neurophysiology and optical imaging studies in monkeys and functional magnetic resonance imaging (fMRI) studies in both monkeys and humans have localized clustered neural responses in inferotemporal cortex selective for images of biologically relevant categories, such as faces and limbs. Using higher resolution (1.5 mm voxels) fMRI scanning methods than past studies (3-5 mm voxels), we recently reported a network of multiple face- and limb-selective regions that neighbor one another in human ventral temporal cortex (Weiner and Grill-Spector, Neuroimage, 52(4):1559-1573, 2010) and lateral occipitotemporal cortex (Weiner and Grill-Spector, Neuroimage, 56(4):2183-2199, 2011). Here, we expand on three basic organization principles of high-level visual cortex revealed by these findings: (1) consistency in the anatomical location of functional regions, (2) preserved spatial relationship among functional regions, and (3) a topographic organization of face- and limb-selective regions in adjacent and alternating clusters. We highlight the implications of this structure in comparing functional brain organization between typical and atypical populations. We conclude with a new model of high-level visual cortex consisting of ventral, lateral, and dorsal components, where multimodal processing related to vision, action, haptics, and language converges in the lateral pathway.
Figures
References
-
- Allison T, Ginter H, McCarthy G, Nobre AC, Puce A, Luby M, et al. Face recognition in human extrastriate cortex. Journal of Neurophysiology. 1994;71(2):821–825. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
