

Gene dose influences cellular and calcium channel dysregulation in heterozygous and homozygous T4826I-RYR1 malignant hyperthermia-susceptible muscle
- PMID: 22139840
- PMCID: PMC3267780
- DOI: 10.1074/jbc.M111.307926
Gene dose influences cellular and calcium channel dysregulation in heterozygous and homozygous T4826I-RYR1 malignant hyperthermia-susceptible muscle
Abstract
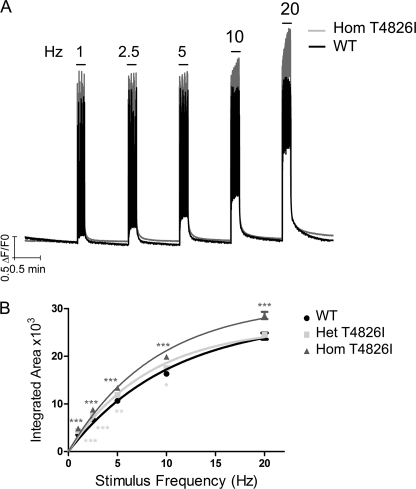
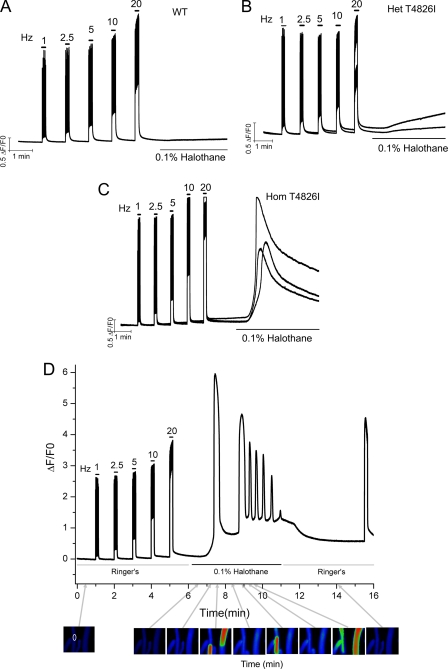
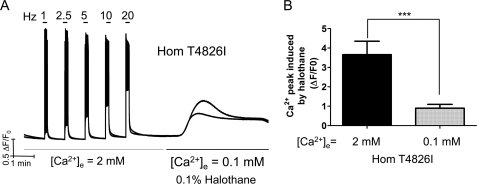
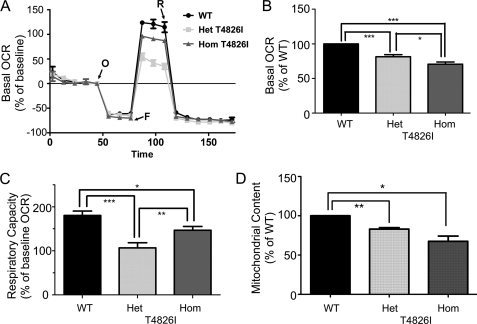
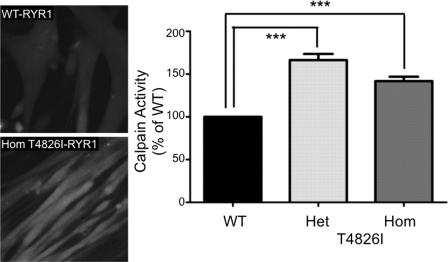
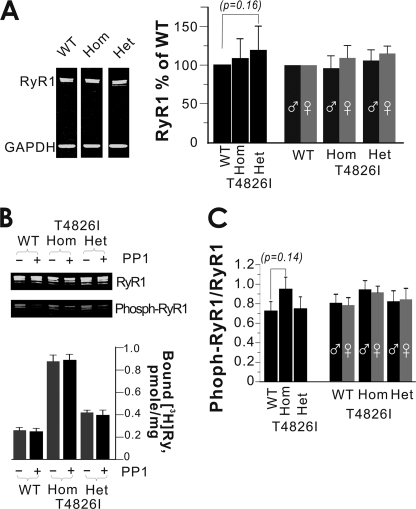
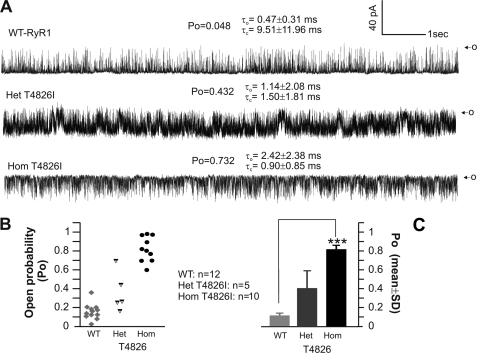
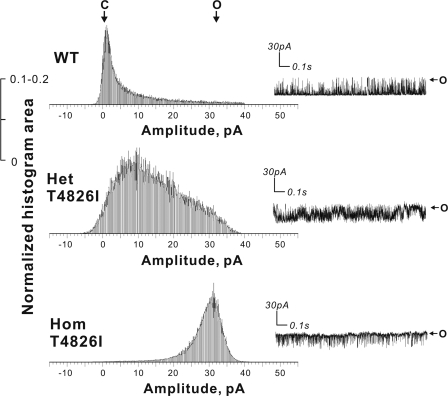
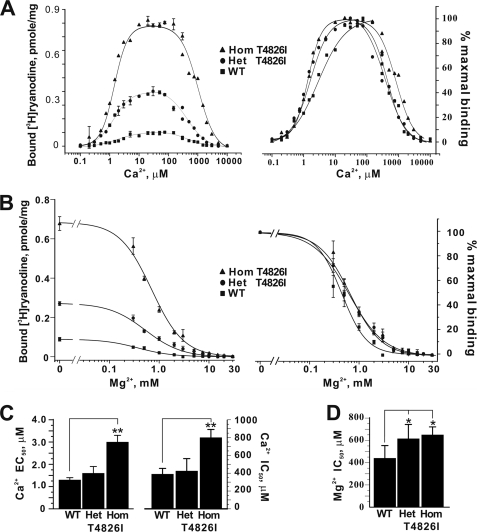
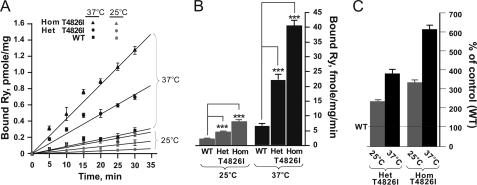
Malignant hyperthermia susceptibility (MHS) is primarily conferred by mutations within ryanodine receptor type 1 (RYR1). Here we address how the MHS mutation T4826I within the S4-S5 linker influences excitation-contraction coupling and resting myoplasmic Ca(2+) concentration ([Ca(2+)](rest)) in flexor digitorum brevis (FDB) and vastus lateralis prepared from heterozygous (Het) and homozygous (Hom) T4826I-RYR1 knock-in mice (Yuen, B. T., Boncompagni, S., Feng, W., Yang, T., Lopez, J. R., Matthaei, K. I., Goth, S. R., Protasi, F., Franzini-Armstrong, C., Allen, P. D., and Pessah, I. N. (2011) FASEB J. doi:22131268). FDB responses to electrical stimuli and acute halothane (0.1%, v/v) exposure showed a rank order of Hom ≫ Het ≫ WT. Release of Ca(2+) from the sarcoplasmic reticulum and Ca(2+) entry contributed to halothane-triggered increases in [Ca(2+)](rest) in Hom FDBs and elicited pronounced Ca(2+) oscillations in ∼30% of FDBs tested. Genotype contributed significantly elevated [Ca(2+)](rest) (Hom > Het > WT) measured in vivo using ion-selective microelectrodes. Het and Hom oxygen consumption rates measured in intact myotubes using the Seahorse Bioscience (Billerica, MA) flux analyzer and mitochondrial content measured with MitoTracker were lower than WT, whereas total cellular calpain activity was higher than WT. Muscle membranes did not differ in RYR1 expression nor in Ser(2844) phosphorylation among the genotypes. Single channel analysis showed highly divergent gating behavior with Hom and WT favoring open and closed states, respectively, whereas Het exhibited heterogeneous gating behaviors. [(3)H]Ryanodine binding analysis revealed a gene dose influence on binding density and regulation by Ca(2+), Mg(2+), and temperature. Pronounced abnormalities inherent in T4826I-RYR1 channels confer MHS and promote basal disturbances of excitation-contraction coupling, [Ca(2+)](rest), and oxygen consumption rates. Considering that both Het and Hom T4826I-RYR1 mice are viable, the remarkable isolated single channel dysfunction mediated through this mutation in S4-S5 cytoplasmic linker must be highly regulated in vivo.
Figures
References
-
- Yuen B. T., Boncompagni S., Feng W., Yang T., Lopez J. R., Matthaei K. I., Goth S. R., Protasi F., Franzini-Armstrong C., Allen P. D., Pessah I. N. (2011) Mice expressing T4826I-RYR1 are viable but exhibit sex- and genotype-dependent susceptibility to malignant hyperthermia and muscle damage. FASEB J. doi:22131268 - PMC - PubMed
-
- Zhou J., Allen P. D., Pessah I. N., Naguib M. (2010) Neuromuscular Disorders and Malignant Hyperthermia. in Miller's Anesthesia (Miller R. D., ed) pp. 1171–1196, 7th Ed., Churchill Livingstone, Philadelphia, PA
-
- Litman R. S., Rosenberg H. (2005) Malignant hyperthermia. Update on susceptibility testing. JAMA 293, 2918–2924 - PubMed
-
- Robinson R., Carpenter D., Shaw M. A., Halsall J., Hopkins P. (2006) Mutations in RYR1 in malignant hyperthermia and central core disease. Hum. Mutat. 27, 977–989 - PubMed
-
- Brady J. E., Sun L. S., Rosenberg H., Li G. (2009) Prevalence of malignant hyperthermia due to anesthesia in New York State, 2001–2005. Anesth. Analg. 109, 1162–1166 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
