

Non-coding telomeric and subtelomeric transcripts are differentially regulated by telomeric and heterochromatin assembly factors in fission yeast
- PMID: 22139922
- PMCID: PMC3326310
- DOI: 10.1093/nar/gkr1155
Non-coding telomeric and subtelomeric transcripts are differentially regulated by telomeric and heterochromatin assembly factors in fission yeast
Abstract
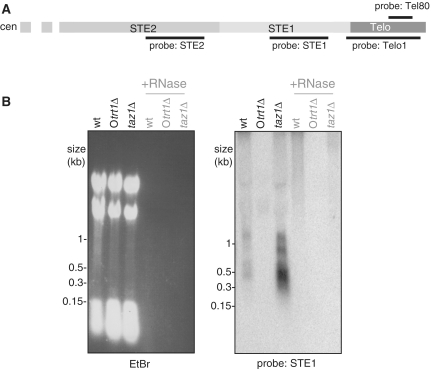
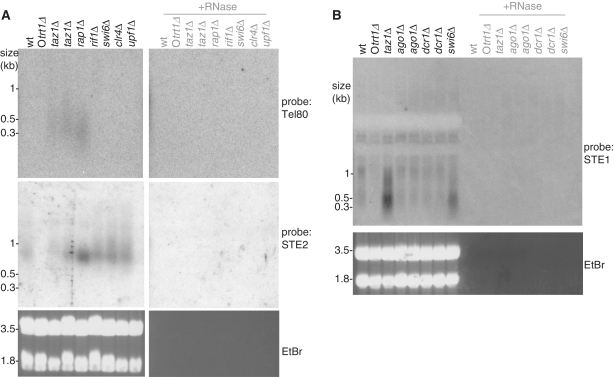
While telomere repeat-containing non-coding RNA has been identified in a variety of eukaryotes, its biological role is not yet clear. We have identified telomeric transcripts in fission yeast, a model system that combines precise genetic manipulability with telomeres remarkably similar to those of human. Like human and budding yeast, fission yeast harbours a population of telomeric RNA molecules containing G-rich telomeric repeats transcribed from the subtelomere to the telomere. In addition, we detect substantial levels of C-rich telomeric RNA whose appearance is independent of the RNA-dependent RNA polymerase, suggesting that the telomere repeats themselves serve as promoter sites; multiple distinct subtelomeric RNAs are also present. The regulation of these transcripts depends on the telomere-associated proteins Taz1 and Rap1, as deletion of taz1(+) or rap1(+) leads to increased levels of both telomere repeat-containing and subtelomeric transcripts. In contrast, loss of the heterochromatin proteins Swi6 or Clr4 or the telomerase regulator Rif1 results in elevated subtelomeric RNA levels while telomere-repeat containing transcript levels remain repressed. Coupled with the large body of knowledge surrounding the functions of telomeric and heterochromatin factors in fission yeast, these in vivo analyses suggest testable models for the roles of TERRA in telomere function.
Figures


Comment on
- Nucleic Acids Res. doi: 10.1093/nar/gkr1153
References
-
- Azzalin CM, Reichenbach P, Khoriauli L, Giulotto E, Lingner J. Telomeric repeat containing RNA and RNA surveillance factors at mammalian chromosome ends. Science. 2007;318:798–801. - PubMed
-
- Schoeftner S, Blasco MA. Developmentally regulated transcription of mammalian telomeres by DNA-dependent RNA polymerase II. Nat. Cell Biol. 2008;10:228–236. - PubMed
-
- Luke B, Panza A, Redon S, Iglesias N, Li Z, Lingner J. The Rat1p 5′ to 3′ exonuclease degrades telomeric repeat-containing RNA and promotes telomere elongation in Saccharomyces cerevisiae. Mol. Cell. 2008;32:465–477. - PubMed
-
- Solovei I, Gaginskaya ER, Macgregor HC. The arrangement and transcription of telomere DNA sequences at the ends of lampbrush chromosomes of birds. Chromosome Res. 1994;2:460–470. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
