

Altered insulin receptor signalling and β-cell cycle dynamics in type 2 diabetes mellitus
- PMID: 22140505
- PMCID: PMC3227614
- DOI: 10.1371/journal.pone.0028050
Altered insulin receptor signalling and β-cell cycle dynamics in type 2 diabetes mellitus
Abstract
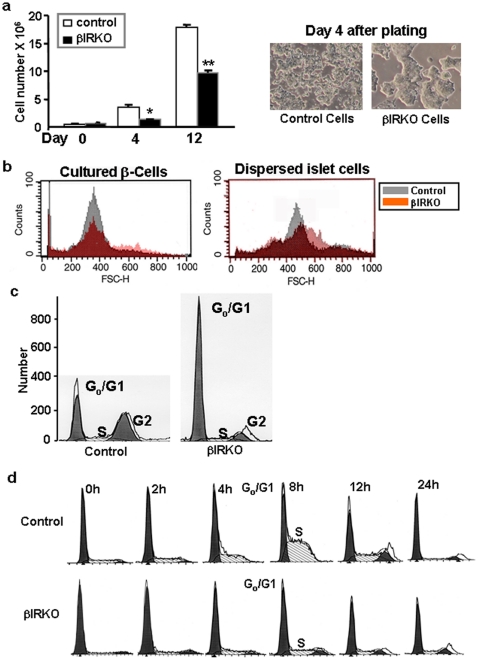
Insulin resistance, reduced β-cell mass, and hyperglucagonemia are consistent features in type 2 diabetes mellitus (T2DM). We used pancreas and islets from humans with T2DM to examine the regulation of insulin signaling and cell-cycle control of islet cells. We observed reduced β-cell mass and increased α-cell mass in the Type 2 diabetic pancreas. Confocal microscopy, real-time PCR and western blotting analyses revealed increased expression of PCNA and down-regulation of p27-Kip1 and altered expression of insulin receptors, insulin receptor substrate-2 and phosphorylated BAD. To investigate the mechanisms underlying these findings, we examined a mouse model of insulin resistance in β-cells--which also exhibits reduced β-cell mass, the β-cell-specific insulin receptor knockout (βIRKO). Freshly isolated islets and β-cell lines derived from βIRKO mice exhibited poor cell-cycle progression, nuclear restriction of FoxO1 and reduced expression of cell-cycle proteins favoring growth arrest. Re-expression of insulin receptors in βIRKO β-cells reversed the defects and promoted cell cycle progression and proliferation implying a role for insulin-signaling in β-cell growth. These data provide evidence that human β- and α-cells can enter the cell-cycle, but proliferation of β-cells in T2DM fails due to G1-to-S phase arrest secondary to defective insulin signaling. Activation of insulin signaling, FoxO1 and proteins in β-cell-cycle progression are attractive therapeutic targets to enhance β-cell regeneration in the treatment of T2DM.
Conflict of interest statement
Figures




References
-
- Accili D. Lilly lecture 2003: the struggle for mastery in insulin action: from triumvirate to republic. Diabetes. 2004;53:1633–1642. - PubMed
-
- DeFronzo RA. Pathogenesis of type 2 diabetes mellitus. Med Clin North Am. 2004;88:787–835. - PubMed
-
- Saltiel AR, Kahn CR. Insulin signalling and the regulation of glucose and lipid metabolism. Nature. 2001;414:799–806. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
