

Type I interferon reaction to viral infection in interferon-competent, immortalized cell lines from the African fruit bat Eidolon helvum
- PMID: 22140523
- PMCID: PMC3227611
- DOI: 10.1371/journal.pone.0028131
Type I interferon reaction to viral infection in interferon-competent, immortalized cell lines from the African fruit bat Eidolon helvum
Abstract
Bats harbor several highly pathogenic zoonotic viruses including Rabies, Marburg, and henipaviruses, without overt clinical symptoms in the animals. It has been suspected that bats might have evolved particularly effective mechanisms to suppress viral replication. Here, we investigated interferon (IFN) response, -induction, -secretion and -signaling in epithelial-like cells of the relevant and abundant African fruit bat species, Eidolon helvum (E. helvum). Immortalized cell lines were generated; their potential to induce and react on IFN was confirmed, and biological assays were adapted to application in bat cell cultures, enabling comparison of landmark IFN properties with that of common mammalian cell lines. E. helvum cells were fully capable of reacting to viral and artificial IFN stimuli. E. helvum cells showed highest IFN mRNA induction, highly productive IFN protein secretion, and evidence of efficient IFN stimulated gene induction. In an Alphavirus infection model, O'nyong-nyong virus exhibited strong IFN induction but evaded the IFN response by translational rather than transcriptional shutoff, similar to other Alphavirus infections. These novel IFN-competent cell lines will allow comparative research on zoonotic, bat-borne viruses in order to model mechanisms of viral maintenance and emergence in bat reservoirs.
Conflict of interest statement
Figures

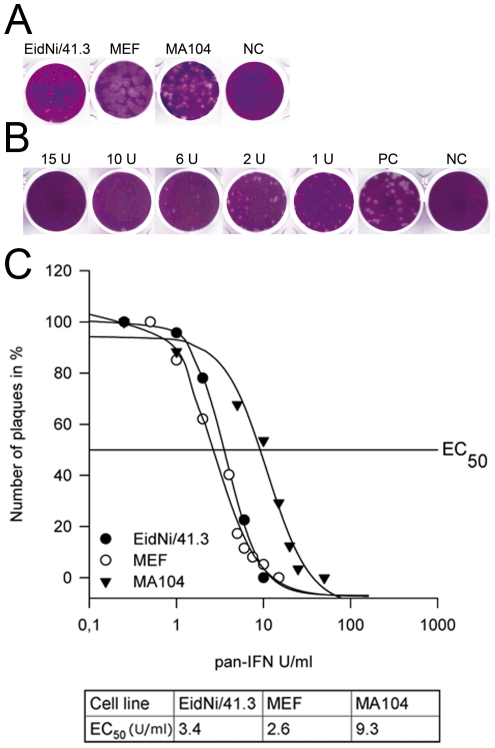

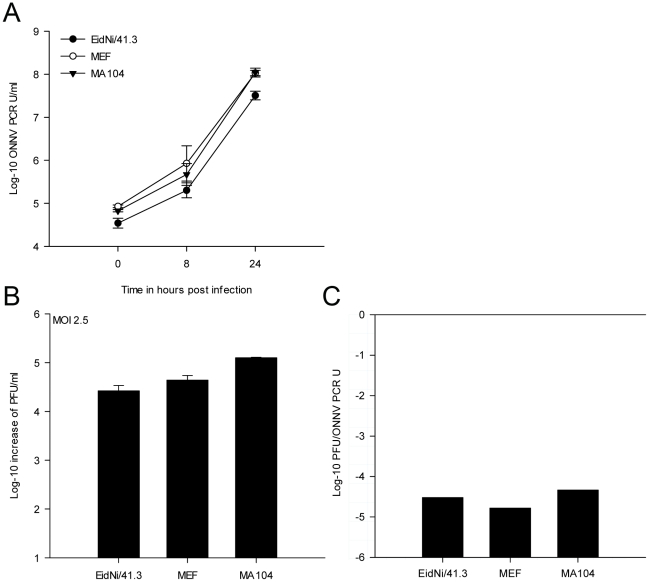
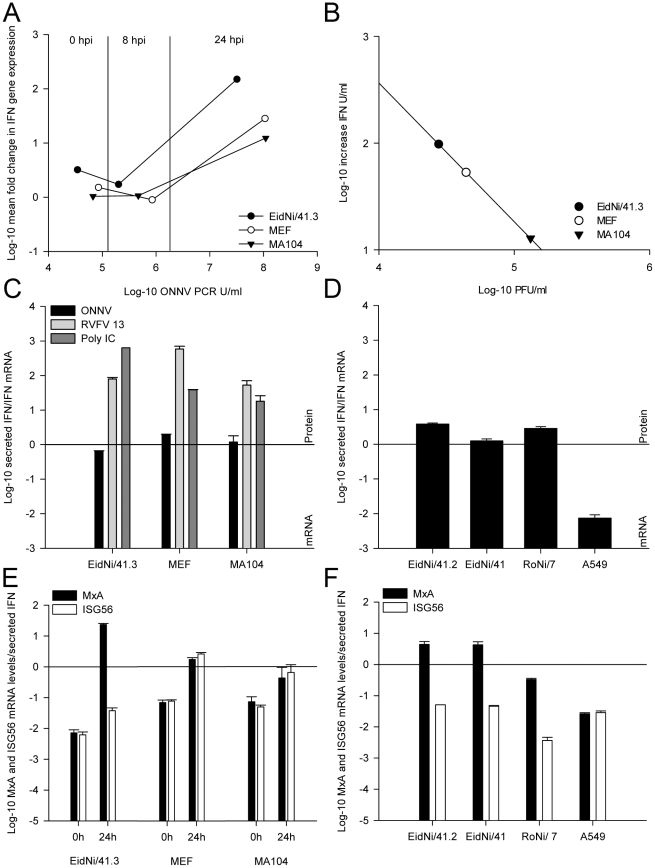
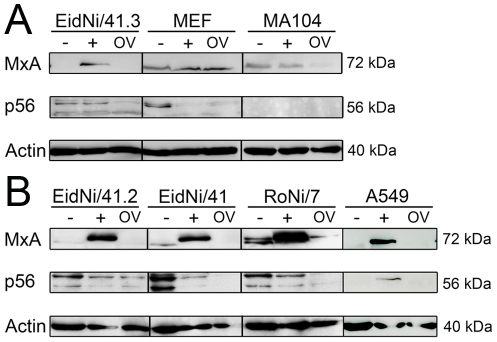

References
-
- Simmons NB. Evolution. An Eocene big bang for bats. Science. 2005;307:527–528. - PubMed
-
- Teeling EC, Springer MS, Madsen O, Bates P, O'Brien S J, et al. A molecular phylogeny for bats illuminates biogeography and the fossil record. Science. 2005;307:580–584. - PubMed
-
- Jones G, Teeling EC. The evolution of echolocation in bats. Trends in Ecology & Evolution. 2006;21:149–156. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
