

Entamoeba lysyl-tRNA synthetase contains a cytokine-like domain with chemokine activity towards human endothelial cells
- PMID: 22140588
- PMCID: PMC3226552
- DOI: 10.1371/journal.pntd.0001398
Entamoeba lysyl-tRNA synthetase contains a cytokine-like domain with chemokine activity towards human endothelial cells
Erratum in
-
Correction: Entamoeba lysyl-tRNA Synthetase Contains a Cytokine-Like Domain with Chemokine Activity towards Human Endothelial Cells.PLoS Negl Trop Dis. 2024 Mar 19;18(3):e0012047. doi: 10.1371/journal.pntd.0012047. eCollection 2024 Mar. PLoS Negl Trop Dis. 2024. PMID: 38502641 Free PMC article.
Abstract
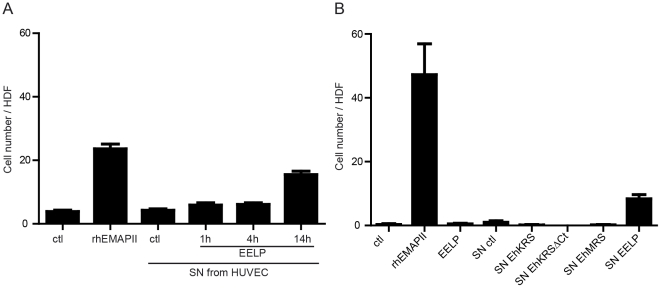
Immunological pressure encountered by protozoan parasites drives the selection of strategies to modulate or avoid the immune responses of their hosts. Here we show that the parasite Entamoeba histolytica has evolved a chemokine that mimics the sequence, structure, and function of the human cytokine HsEMAPII (Homo sapiens endothelial monocyte activating polypeptide II). This Entamoeba EMAPII-like polypeptide (EELP) is translated as a domain attached to two different aminoacyl-tRNA synthetases (aaRS) that are overexpressed when parasites are exposed to inflammatory signals. EELP is dispensable for the tRNA aminoacylation activity of the enzymes that harbor it, and it is cleaved from them by Entamoeba proteases to generate a standalone cytokine. Isolated EELP acts as a chemoattractant for human cells, but its cell specificity is different from that of HsEMAPII. We show that cell specificity differences between HsEMAPII and EELP can be swapped by site directed mutagenesis of only two residues in the cytokines' signal sequence. Thus, Entamoeba has evolved a functional mimic of an aaRS-associated human cytokine with modified cell specificity.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Similar articles
-
Crystal structures of Entamoeba histolytica lysyl-tRNA synthetase reveal conformational changes upon lysine binding and a specific helix bundle domain.FEBS Lett. 2014 Nov 28;588(23):4478-86. doi: 10.1016/j.febslet.2014.10.019. FEBS Lett. 2014. PMID: 25448989
-
Structure of the EMAPII domain of human aminoacyl-tRNA synthetase complex reveals evolutionary dimer mimicry.EMBO J. 2001 Feb 1;20(3):570-8. doi: 10.1093/emboj/20.3.570. EMBO J. 2001. PMID: 11157763 Free PMC article.
-
Unusual domain architecture of aminoacyl tRNA synthetases and their paralogs from Leishmania major.BMC Genomics. 2012 Nov 14;13:621. doi: 10.1186/1471-2164-13-621. BMC Genomics. 2012. PMID: 23151081 Free PMC article.
-
Lysyl-tRNA synthetase.Biol Chem Hoppe Seyler. 1995 Aug;376(8):451-72. doi: 10.1515/bchm3.1995.376.8.451. Biol Chem Hoppe Seyler. 1995. PMID: 7576245 Review.
-
Structural analyses of the malaria parasite aminoacyl-tRNA synthetases provide new avenues for antimalarial drug discovery.Protein Sci. 2021 Sep;30(9):1793-1803. doi: 10.1002/pro.4148. Protein Sci. 2021. PMID: 34184352 Free PMC article. Review.
Cited by
-
Genetic Validation of Leishmania donovani Lysyl-tRNA Synthetase Shows that It Is Indispensable for Parasite Growth and Infectivity.mSphere. 2017 Aug 30;2(4):e00340-17. doi: 10.1128/mSphereDirect.00340-17. eCollection 2017 Jul-Aug. mSphere. 2017. PMID: 28875178 Free PMC article.
-
The Aeromonas salmonicida subsp. salmonicida exoproteome: global analysis, moonlighting proteins and putative antigens for vaccination against furunculosis.Proteome Sci. 2013 Oct 15;11(1):44. doi: 10.1186/1477-5956-11-44. Proteome Sci. 2013. PMID: 24127837 Free PMC article.
-
Correction: Entamoeba lysyl-tRNA Synthetase Contains a Cytokine-Like Domain with Chemokine Activity towards Human Endothelial Cells.PLoS Negl Trop Dis. 2024 Mar 19;18(3):e0012047. doi: 10.1371/journal.pntd.0012047. eCollection 2024 Mar. PLoS Negl Trop Dis. 2024. PMID: 38502641 Free PMC article.
-
tRNA synthetase: tRNA aminoacylation and beyond.Wiley Interdiscip Rev RNA. 2014 Jul-Aug;5(4):461-80. doi: 10.1002/wrna.1224. Epub 2014 Apr 4. Wiley Interdiscip Rev RNA. 2014. PMID: 24706556 Free PMC article. Review.
-
Twin Attributes of Tyrosyl-tRNA Synthetase of Leishmania donovani: A HOUSEKEEPING PROTEIN TRANSLATION ENZYME AND A MIMIC OF HOST CHEMOKINE.J Biol Chem. 2016 Aug 19;291(34):17754-71. doi: 10.1074/jbc.M116.727107. Epub 2016 Jul 5. J Biol Chem. 2016. PMID: 27382051 Free PMC article.
References
-
- Ackers JP, Mirelman D. Progress in research on Entamoeba histolytica pathogenesis. Curr Opin Microbiol. 2006;9:367–373. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
