

A hybrid BAC physical map of potato: a framework for sequencing a heterozygous genome
- PMID: 22142254
- PMCID: PMC3261212
- DOI: 10.1186/1471-2164-12-594
A hybrid BAC physical map of potato: a framework for sequencing a heterozygous genome
Erratum in
- BMC Genomics. 2012;13:423. Wiggers-Perebolte, Lian [added]; de Leeuw, Linda [added]
Abstract
Background: Potato is the world's third most important food crop, yet cultivar improvement and genomic research in general remain difficult because of the heterozygous and tetraploid nature of its genome. The development of physical map resources that can facilitate genomic analyses in potato has so far been very limited. Here we present the methods of construction and the general statistics of the first two genome-wide BAC physical maps of potato, which were made from the heterozygous diploid clone RH89-039-16 (RH).
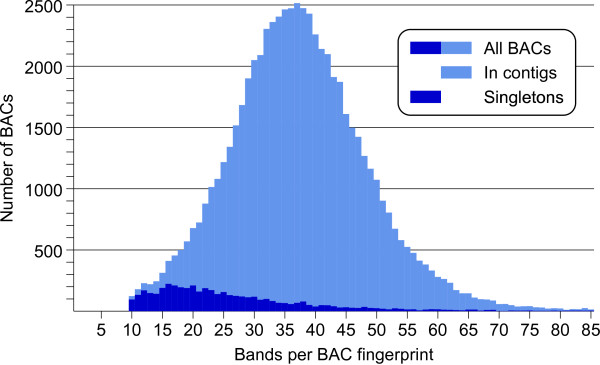
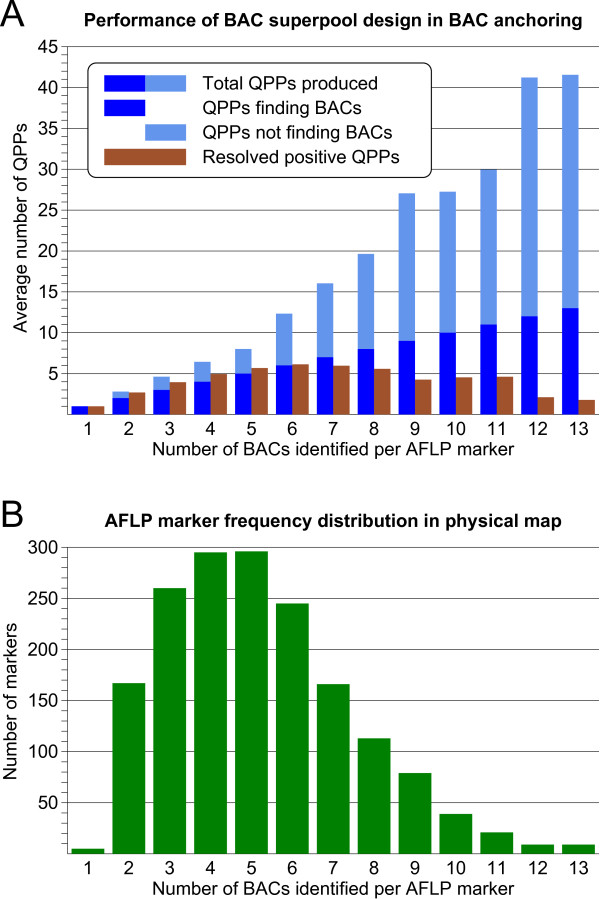
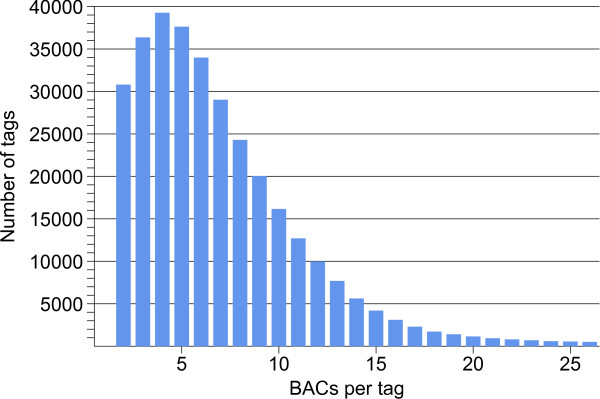
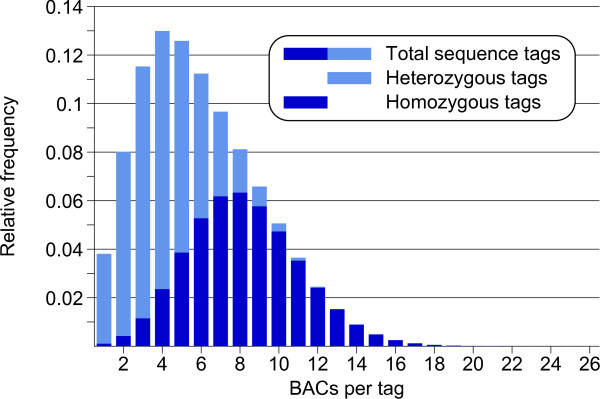
Results: First, a gel electrophoresis-based physical map was made by AFLP fingerprinting of 64478 BAC clones, which were aligned into 4150 contigs with an estimated total length of 1361 Mb. Screening of BAC pools, followed by the KeyMaps in silico anchoring procedure, identified 1725 AFLP markers in the physical map, and 1252 BAC contigs were anchored the ultradense potato genetic map. A second, sequence-tag-based physical map was constructed from 65919 whole genome profiling (WGP) BAC fingerprints and these were aligned into 3601 BAC contigs spanning 1396 Mb. The 39733 BAC clones that overlap between both physical maps provided anchors to 1127 contigs in the WGP physical map, and reduced the number of contigs to around 2800 in each map separately. Both physical maps were 1.64 times longer than the 850 Mb potato genome. Genome heterozygosity and incomplete merging of BAC contigs are two factors that can explain this map inflation. The contig information of both physical maps was united in a single table that describes hybrid potato physical map.
Conclusions: The AFLP physical map has already been used by the Potato Genome Sequencing Consortium for sequencing 10% of the heterozygous genome of clone RH on a BAC-by-BAC basis. By layering a new WGP physical map on top of the AFLP physical map, a genetically anchored genome-wide framework of 322434 sequence tags has been created. This reference framework can be used for anchoring and ordering of genomic sequences of clone RH (and other potato genotypes), and opens the possibility to finish sequencing of the RH genome in a more efficient way via high throughput next generation approaches.
Figures







Similar articles
-
A high-throughput AFLP-based method for constructing integrated genetic and physical maps: progress toward a sorghum genome map.Genome Res. 2000 Jun;10(6):789-807. doi: 10.1101/gr.10.6.789. Genome Res. 2000. PMID: 10854411 Free PMC article.
-
Construction of a 10,000-marker ultradense genetic recombination map of potato: providing a framework for accelerated gene isolation and a genomewide physical map.Genetics. 2006 Jun;173(2):1075-87. doi: 10.1534/genetics.106.055871. Epub 2006 Apr 2. Genetics. 2006. PMID: 16582432 Free PMC article.
-
Exploitation of a marker dense linkage map of potato for positional cloning of a wart disease resistance gene.Theor Appl Genet. 2006 Jan;112(2):269-77. doi: 10.1007/s00122-005-0125-x. Epub 2005 Oct 29. Theor Appl Genet. 2006. PMID: 16261329
-
Advances in BAC-based physical mapping and map integration strategies in plants.J Biomed Biotechnol. 2012;2012:184854. doi: 10.1155/2012/184854. Epub 2012 Feb 27. J Biomed Biotechnol. 2012. PMID: 22500080 Free PMC article. Review.
-
Generation of physical map contig-specific sequences.Front Genet. 2014 Jul 22;5:243. doi: 10.3389/fgene.2014.00243. eCollection 2014. Front Genet. 2014. PMID: 25101119 Free PMC article. Review.
Cited by
-
Whole genome profiling physical map and ancestral annotation of tobacco Hicks Broadleaf.Plant J. 2013 Sep;75(5):880-9. doi: 10.1111/tpj.12247. Epub 2013 Jun 26. Plant J. 2013. PMID: 23672264 Free PMC article.
-
High-density interspecific genetic maps of kiwifruit and the identification of sex-specific markers.DNA Res. 2015 Oct;22(5):367-75. doi: 10.1093/dnares/dsv019. Epub 2015 Sep 14. DNA Res. 2015. PMID: 26370666 Free PMC article.
-
Construction of reference chromosome-scale pseudomolecules for potato: integrating the potato genome with genetic and physical maps.G3 (Bethesda). 2013 Nov 6;3(11):2031-47. doi: 10.1534/g3.113.007153. G3 (Bethesda). 2013. PMID: 24062527 Free PMC article.
-
Potato genetics, genomics, and applications.Breed Sci. 2015 Mar;65(1):53-68. doi: 10.1270/jsbbs.65.53. Epub 2015 Mar 1. Breed Sci. 2015. PMID: 25931980 Free PMC article. Review.
-
Development of a Sequence-Based Reference Physical Map of Pea (Pisum sativum L.).Front Plant Sci. 2019 Mar 15;10:323. doi: 10.3389/fpls.2019.00323. eCollection 2019. Front Plant Sci. 2019. PMID: 30930928 Free PMC article.
References
-
- Arumuganathan K, Earle ED. Nuclear DNA content of some important plant species. Plant Mol Biol Rep. 1991;9:208–218. doi: 10.1007/BF02672069. - DOI
-
- Bradshaw JE, Pande B, Bryan GJ, Hackett CA, McLean K, Stewart HE, Waugh R. Interval mapping of quantitative trait loci for resistance to late blight [Phytophthora infestans (Mont.) de Bary], height and maturity in a tetraploid population of potato (Solanum tuberosum subsp. tuberosum) Genetics. 2004;168:983–995. doi: 10.1534/genetics.104.030056. - DOI - PMC - PubMed
-
- Carputo D, Barone A. Ploidy level manipulations in potato through sexual hybridisation. Ann Appl Biol. 2005;146:71–79. doi: 10.1111/j.1744-7348.2005.04070.x. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
