

Sterol-induced degradation of HMG CoA reductase depends on interplay of two Insigs and two ubiquitin ligases, gp78 and Trc8
- PMID: 22143767
- PMCID: PMC3251157
- DOI: 10.1073/pnas.1112831108
Sterol-induced degradation of HMG CoA reductase depends on interplay of two Insigs and two ubiquitin ligases, gp78 and Trc8
Abstract
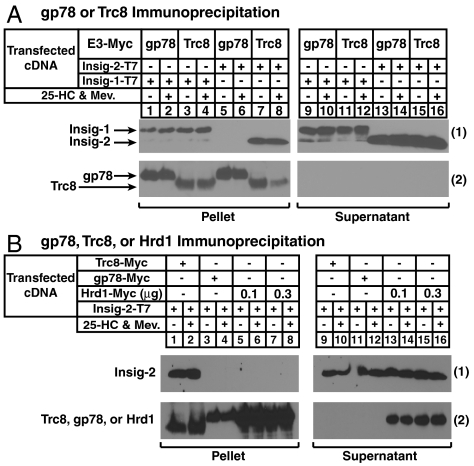
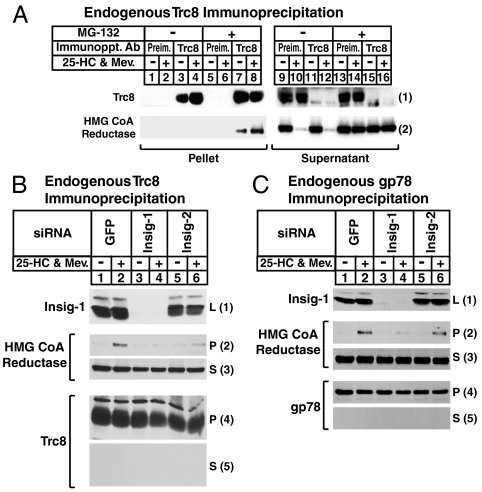
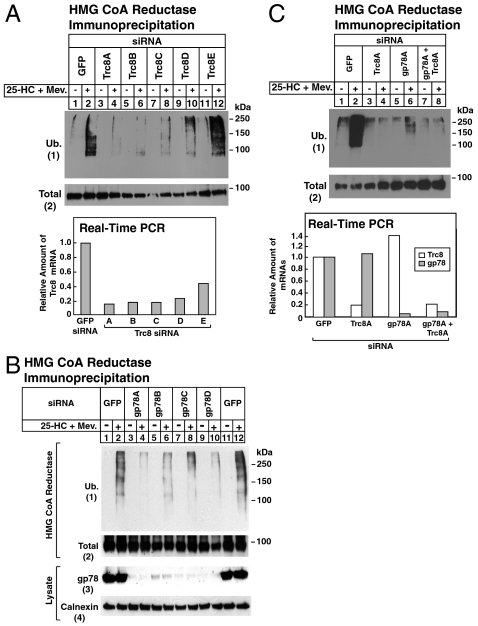
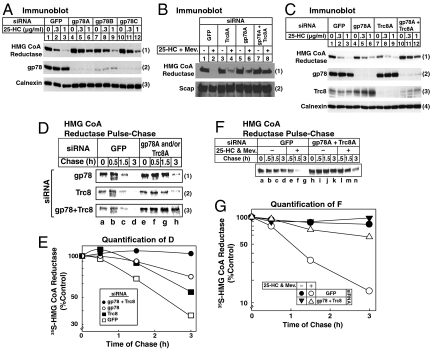
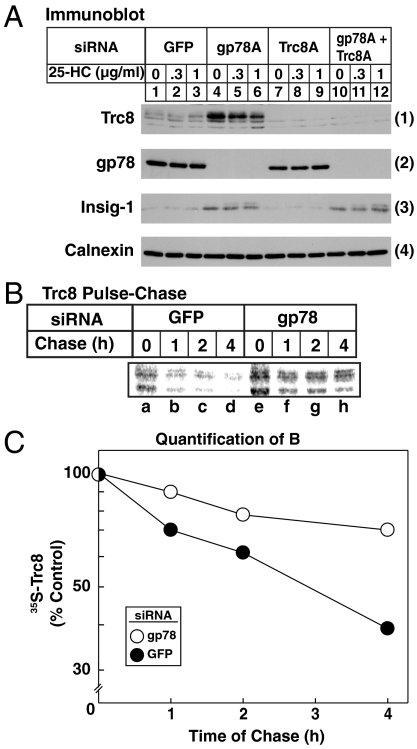
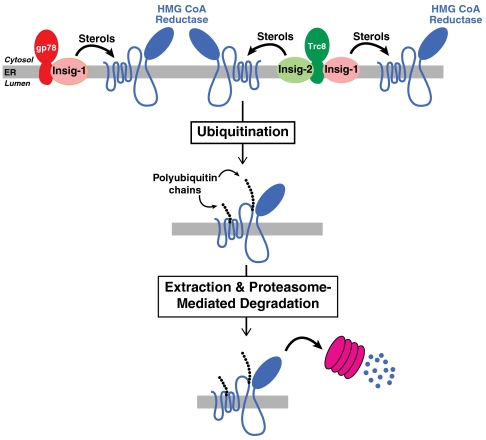
Accumulation of sterols in membranes of the endoplasmic reticulum (ER) leads to the accelerated ubiquitination and proteasomal degradation of 3-hydroxy-3-methylglutaryl coenzyme A reductase, a rate-limiting enzyme in synthesis of cholesterol and nonsterol isoprenoids. This degradation results from sterol-induced binding of reductase to the Insig-1 or Insig-2 proteins of ER membranes. We previously reported that in immortalized human fibroblasts (SV-589 cells) Insig-1, but not Insig-2, recruits gp78, a membrane-bound RING-finger ubiquitin ligase. We now report that both Insig-1 and Insig-2 bind another membrane-bound RING-finger ubiquitin ligase called Trc8. Knockdown of either gp78 or Trc8 in SV-589 cells through RNA interference (RNAi) inhibited sterol-induced ubiquitination of reductase and inhibited sterol-induced degradation by 50-60%. The combined knockdown of gp78 and Trc8 produced a more complete inhibition of degradation (> 90%). Knockdown of gp78 led to a three to fourfold increase in levels of Trc8 and Insig-1 proteins, which opposed the inhibitory action of gp78. In contrast, knockdown of Trc8 had no effect on gp78 or Insig-1. The current results suggest that sterol-induced ubiquitination and proteasomal degradation of reductase is dictated by the complex interplay of at least four proteins: Insig-1, Insig-2, gp78, and Trc8. Variations in the concentrations of any one of these proteins may account for differences in cell- and/or tissue-specific regulation of reductase degradation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Sterol-induced dislocation of 3-hydroxy-3-methylglutaryl coenzyme A reductase from membranes of permeabilized cells.Mol Biol Cell. 2013 Nov;24(21):3300-8. doi: 10.1091/mbc.E13-03-0157. Epub 2013 Sep 11. Mol Biol Cell. 2013. PMID: 24025715 Free PMC article.
-
Ring finger protein 145 (RNF145) is a ubiquitin ligase for sterol-induced degradation of HMG-CoA reductase.J Biol Chem. 2018 Mar 16;293(11):4047-4055. doi: 10.1074/jbc.RA117.001260. Epub 2018 Jan 26. J Biol Chem. 2018. PMID: 29374057 Free PMC article.
-
Ancient ubiquitous protein-1 mediates sterol-induced ubiquitination of 3-hydroxy-3-methylglutaryl CoA reductase in lipid droplet-associated endoplasmic reticulum membranes.Mol Biol Cell. 2013 Feb;24(3):169-83. doi: 10.1091/mbc.E12-07-0564. Epub 2012 Dec 5. Mol Biol Cell. 2013. PMID: 23223569 Free PMC article.
-
Control of cholesterol synthesis through regulated ER-associated degradation of HMG CoA reductase.Crit Rev Biochem Mol Biol. 2010 Jun;45(3):185-98. doi: 10.3109/10409238.2010.485605. Crit Rev Biochem Mol Biol. 2010. PMID: 20482385 Free PMC article. Review.
-
Feedback regulation of cholesterol synthesis: sterol-accelerated ubiquitination and degradation of HMG CoA reductase.Cell Res. 2008 Jun;18(6):609-21. doi: 10.1038/cr.2008.61. Cell Res. 2008. PMID: 18504457 Free PMC article. Review.
Cited by
-
Vitamin D supplementation is effective for olanzapine-induced dyslipidemia.Front Pharmacol. 2023 Feb 21;14:1135516. doi: 10.3389/fphar.2023.1135516. eCollection 2023. Front Pharmacol. 2023. PMID: 36895943 Free PMC article.
-
Sterol-induced dislocation of 3-hydroxy-3-methylglutaryl coenzyme A reductase from membranes of permeabilized cells.Mol Biol Cell. 2013 Nov;24(21):3300-8. doi: 10.1091/mbc.E13-03-0157. Epub 2013 Sep 11. Mol Biol Cell. 2013. PMID: 24025715 Free PMC article.
-
Type 1 polyisoprenoid diphosphate phosphatase modulates geranylgeranyl-mediated control of HMG CoA reductase and UBIAD1.Elife. 2021 Nov 29;10:e64688. doi: 10.7554/eLife.64688. Elife. 2021. PMID: 34842525 Free PMC article.
-
The E3 ubiquitin ligases, HUWE1 and NEDD4-1, are involved in the post-translational regulation of the ABCG1 and ABCG4 lipid transporters.J Biol Chem. 2015 Oct 2;290(40):24604-13. doi: 10.1074/jbc.M115.675579. Epub 2015 Aug 20. J Biol Chem. 2015. PMID: 26296893 Free PMC article.
-
Hypoxia-inducible factor 1α activates insulin-induced gene 2 (Insig-2) transcription for degradation of 3-hydroxy-3-methylglutaryl (HMG)-CoA reductase in the liver.J Biol Chem. 2017 Jun 2;292(22):9382-9393. doi: 10.1074/jbc.M117.788562. Epub 2017 Apr 17. J Biol Chem. 2017. PMID: 28416613 Free PMC article.
References
-
- Brown MS, Goldstein JL. Multivalent feedback regulation of HMG CoA reductase, a control mechanism coordinating isoprenoid synthesis and cell growth. J Lipid Res. 1980;21:505–517. - PubMed
-
- Sever N, Yang T, Brown MS, Goldstein JL, DeBose-Boyd RA. Accelerated degradation of HMG CoA reductase mediated by binding of insig-1 to its sterol-sensing domain. Mol Cell. 2003;11:25–33. - PubMed
-
- Sever N, et al. Insig-dependent ubiquitination and degradation of mammalian 3-hydroxy-3-methylglutaryl-CoA reductase stimulated by sterols and geranylgeraniol. J Biol Chem. 2003;278:52479–52490. - PubMed
-
- Liscum L, et al. Domain structure of 3-hydroxy-3-methylglutaryl coenzyme A reductase, a glycoprotein of the endoplasmic reticulum. J Biol Chem. 1985;260:522–530. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
