

Paraquat neurotoxicity is mediated by the dopamine transporter and organic cation transporter-3
- PMID: 22143804
- PMCID: PMC3251116
- DOI: 10.1073/pnas.1115141108
Paraquat neurotoxicity is mediated by the dopamine transporter and organic cation transporter-3
Abstract
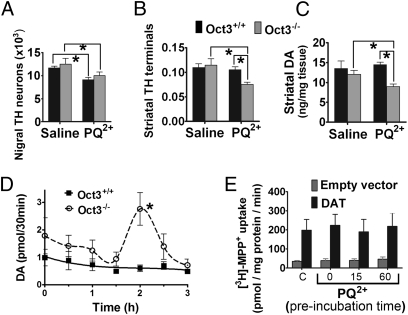
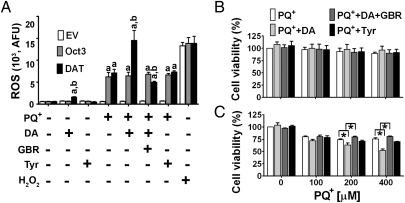
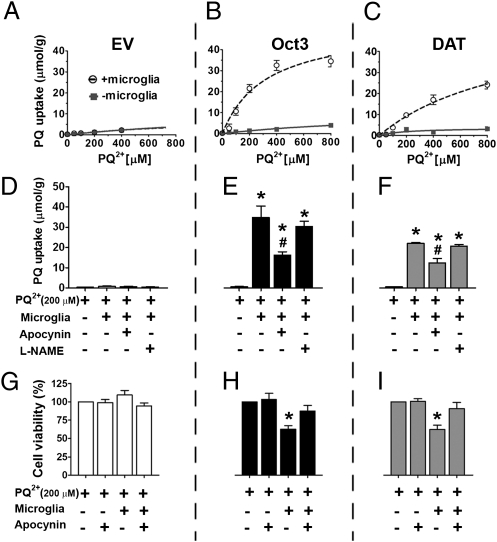
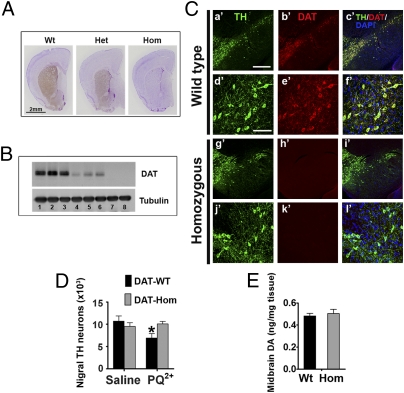
The herbicide paraquat (PQ) has increasingly been reported in epidemiological studies to enhance the risk of developing Parkinson's disease (PD). Furthermore, case-control studies report that individuals with genetic variants in the dopamine transporter (DAT, SLC6A) have a higher PD risk when exposed to PQ. However, it remains a topic of debate whether PQ can enter dopamine (DA) neurons through DAT. We report here a mechanism by which PQ is transported by DAT: In its native divalent cation state, PQ(2+) is not a substrate for DAT; however, when converted to the monovalent cation PQ(+) by either a reducing agent or NADPH oxidase on microglia, it becomes a substrate for DAT and is accumulated in DA neurons, where it induces oxidative stress and cytotoxicity. Impaired DAT function in cultured cells and mutant mice significantly attenuated neurotoxicity induced by PQ(+). In addition to DAT, PQ(+) is also a substrate for the organic cation transporter 3 (Oct3, Slc22a3), which is abundantly expressed in non-DA cells in the nigrostriatal regions. In mice with Oct3 deficiency, enhanced striatal damage was detected after PQ treatment. This increased sensitivity likely results from reduced buffering capacity by non-DA cells, leading to more PQ(+) being available for uptake by DA neurons. This study provides a mechanism by which DAT and Oct3 modulate nigrostriatal damage induced by PQ(2+)/PQ(+) redox cycling.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
- T32 GM07356/GM/NIGMS NIH HHS/United States
- ES020081/ES/NIEHS NIH HHS/United States
- R21 ES017470/ES/NIEHS NIH HHS/United States
- ES17470/ES/NIEHS NIH HHS/United States
- TL1RR 024135/RR/NCRR NIH HHS/United States
- UL1 RR024160/RR/NCRR NIH HHS/United States
- ES0017470-01S1/ES/NIEHS NIH HHS/United States
- F30 AG040903/AG/NIA NIH HHS/United States
- F30 ES020081/ES/NIEHS NIH HHS/United States
- DA022413/DA/NIDA NIH HHS/United States
- T32 GM007356/GM/NIGMS NIH HHS/United States
- ES014899/ES/NIEHS NIH HHS/United States
- TL1 RR024135/RR/NCRR NIH HHS/United States
- K05 DA022413/DA/NIDA NIH HHS/United States
- R01 ES014899/ES/NIEHS NIH HHS/United States
- AG040903/AG/NIA NIH HHS/United States
- P01 DA012408/DA/NIDA NIH HHS/United States
- DA12408/DA/NIDA NIH HHS/United States
- KL2 RR024136/RR/NCRR NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
